La retina es la porción del ojo sensible a la luz que contiene, los conos que responsables de la visión de los colores y los bastones que detectan luz tenue y están encargados básicamente de la visión en blanco y negro y de la visión en la oscuridad.
Las capas de la retina desde el exterior hacia el interior:
1) capa pigmentaria
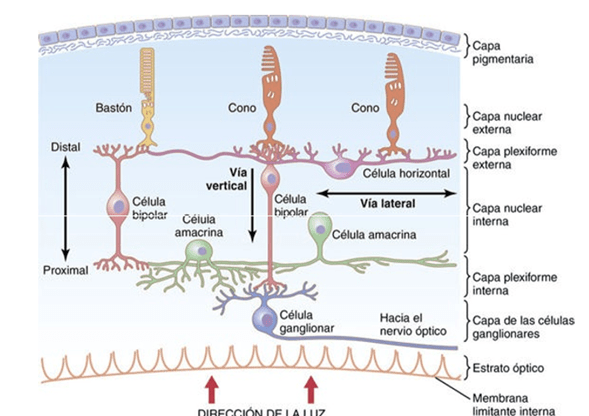
2) capa de conos y bastones que aloja las prolongaciones de estos receptores hacia la capa anterior
3) capa nuclear externa que contiene los somas de los conos y los bastones
4) capa plexiforme externa
5) capa nuclear interna
6) capa plexiforme interna
7) capa ganglionar
8) capa de las fibras del nervio óptico
9) membrana limitante interna.
La región de la fóvea de la retina es una zona diminuta en el centro de la retina, que ocupa un área total un poco mayor de 1 mm, se encuentra especialmente capacitada para la visión aguda y detallada. La fóvea central, de solo 0,3 mm de diámetro, está compuesta por conos casi en su integridad. posee una estructura especial que contribuye a la detección de detalles en la imagen visual, los conos de la fóvea presentan un cuerpo especialmente largo y delgado, en contraposición a los conos situados hacia la periferia de la retina, que son mucho más gruesos.

Los conos y bastones, es una representación esquemática de los componentes esenciales de un fotorreceptor. El segmento externo del cono posee una forma cónica. En las porciones periféricas de la retina, los bastones alcanzan un diámetro de 2 a 5 μm, mientras que los conos miden de 5 a 8; en la parte central de la retina, la fóvea, hay bastones, y los conos son delgados y su diámetro solo es de 1,5 μm. En el caso de los bastones, esta sustancia fotosensible es la rodopsina; en los conos, es una de las tres sustancias fotosensibles al «color», que suelen denominarse simplemente pigmentos del color, y funcionan casi exactamente igual que la rodopsina excepto por sus diferencias de sensibilidad dentro del espectro. En cada cono o bastón llega a haber hasta 1.000 discos. Tanto la rodopsina como los pigmentos de color son proteínas conjugadas. Se incorporan a las membranas de los discos bajo la forma de proteínas transmembrana. Las concentraciones de estos pigmentos fotosensibles en los discos son tan grandes que ellos mismos constituyen aproximadamente el 40% de toda la masa del segmento externo. El segmento interno del cono o del bastón contiene el citoplasma habitual con los orgánulos citoplásmicos. Especialmente importantes son las mitocondrias que, según se explica más adelante, cumplen la decisiva misión de proporcionar energía para el funcionamiento de los fotorreceptores.

El potencial de receptor del bastón es hiperpolarizante, no despolarizante, cuando el bastón se encuentra expuesto a la luz, el potencial de receptor resultante es diferente del que existe en casi todos los demás receptores sensitivos, la excitación de un bastón provoca un aumento de la negatividad en el potencial de membrana en su interior, lo que supone un estado de hiperpolarización. es exactamente lo contrario del descenso de la negatividad «despolarización» que sucede en todos los demás receptores sensitivos. Cuando se descompone la rodopsina, disminuye la conductancia de la membrana del bastón para los iones sodio en su segmento externo.
provoca una hiperpolarización de toda la membrana del bastón por el siguiente mecanismo. el movimiento de los iones sodio y potasio en un circuito eléctrico completo que atraviesa los segmentos interno y externo del bastón. El segmento interno bombea sodio sin interrupción desde el interior del bastón hacia su exterior y se bombean iones potasio hacia el interior de la célula. Los iones potasio se filtran de la célula a través de canales de potasio no activados que están confinados en el segmento interno del bastón. bomba de sodio-potasio crea un potencial negativo dentro de la célula tomada en su integridad.
En el estado de oscuridad, los niveles de GMPc son elevados, lo que hace posible que estos iones de carga positiva se filtren permanentemente hacia el interior del bastón y neutralicen así gran parte de la negatividad que hay dentro de toda la célula. Por tanto, en condiciones de oscuridad normales, cuando el bastón no está excitado, hay una baja electronegatividad en la parte interna a su membrana, que mide unos –40 mV en vez de los habituales –70 a –80 presentes en la mayoría de los receptores sensitivos
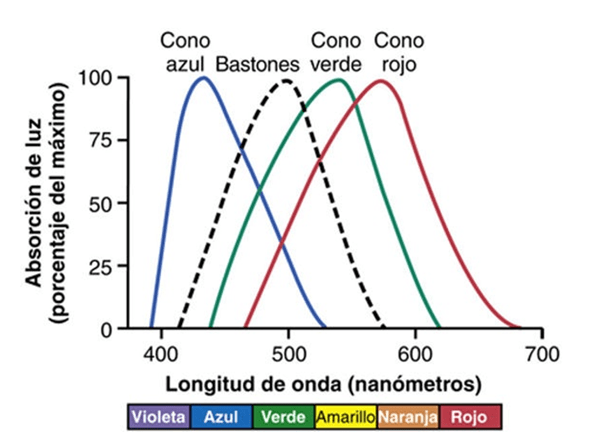
Fotoquímica de la visión de los colores por los conos. La porción de todos los pigmentos visuales correspondiente al retinal es absolutamente idéntica en los conos y en los bastones. los pigmentos sensibles al color de los conos consisten en combinaciones de retinal y fotopsinas. Los diversos colores: azul, verde o rojo. Estos pigmentos para el color se llaman, respectivamente, pigmento sensible al azul, pigmento sensible al verde y pigmento sensible al rojo. Sus características de absorción en los tres tipos de conos muestran unas absorbencias máximas ante la luz de las
longitudes de onda de 445, 535 y 570 nm, respectivamente. Estas son también las longitudes de onda con una sensibilidad extrema a la luz en cada tipo de cono, lo que empieza a sentar las condiciones para poder explicar cómo distingue los colores la retina. También se ofrece la curva de absorción de la rodopsina en los bastones, con un pico a 505 nm.
Percepción de la luz blanca, una estimulación equivalente de los conos rojo, verde y azul da lugar a la sensación visual del blanco. Con todo, la luz no tiene una sola longitud de onda que corresponda al blanco; este color es una combinación de todas las longitudes del espectro. La percepción del blanco puede lograrse mediante la estimulación de la retina con la combinación oportuna tan solo de tres colores escogidos que activen los tipos de conos respectivos más o menos por igual.
La función nerviosa de la retina presenta los puntos fundamentales donde se producen sus conexiones nerviosas, con el circuito de la retina periférica a la izquierda y el circuito de la zona de la fóvea a la derecha.
diversos tipos neuronales son:
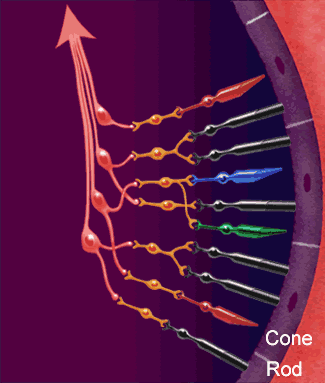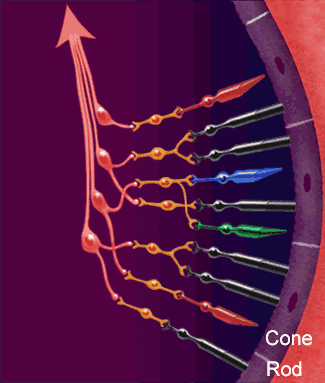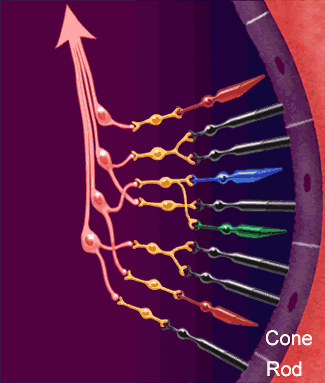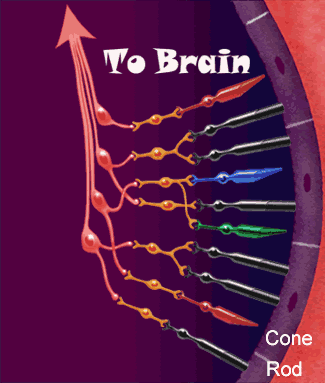
1. Los fotorreceptores, los conos y los bastones, que transmiten las señales hacia la capa plexiforme externa, donde hacen sinapsis con las células bipolares y horizontales.
2. Las células horizontales, que transmiten las señales en sentido horizontal por la capa plexiforme externa desde los conos y los bastones hasta las células bipolares.
3. Las células bipolares, que transmiten las señales en sentido vertical desde los conos, los bastones y las células horizontales hacia la capa plexiforme interna, donde hacen sinapsis con las células ganglionares y amacrinas.
4. Las células amacrinas, que transmiten las señales en dos direcciones, directamente desde las células bipolares hasta las células ganglionares, u horizontalmente en el seno de la capa plexiforme interna desde los axones de las células bipolares hasta las dendritas de las células ganglionares o hasta otras células amacrinas.
5. Las células ganglionares, que transmiten las señales de salida desde la retina hacia el cerebro a través del nervio óptico.
La retina posee un tipo antiguo de visión basado en los bastones y otro nuevo que reposa en los conos. Las neuronas y las fibras nerviosas encargadas de conducir las señales visuales correspondientes a la visión de los conos son considerablemente mayores que las encargadas de la visión de los bastones y los impulsos se envían al cerebro con una velocidad de dos a cinco veces superior.
Los conos y los bastones liberan glutamato en sus sinapsis con las células bipolares. Los estudios histológicos y farmacológicos han demostrado la existencia de muchas clases de células amacrinas que segregan un mínimo de ocho tipos de sustancias transmisoras, como ácido γ aminobutírico (GABA), glicina, dopamina, acetilcolina e indolamina, cuya acción normalmente posee en general un carácter inhibidor. Los de las células bipolares, horizontales e interplexiformes no están nada claros, pero al menos parte de las células horizontales liberan transmisores inhibidores.

Potenciales de acción continuos y espontáneos
Las células ganglionares son el punto de origen de las fibras largas que llegan al cerebro formando el nervio óptico. Dada la distancia que han de recorrer, el método de conducción electrotónico empleado por los conos, los bastones y las células bipolares en el interior de la retina deja de ser rentable; por tanto, las células ganglionares pasan a transmitir sus impulsos mediante potenciales de acción repetidos. Además, incluso cuando no están estimuladas, envían impulsos continuos a una frecuencia que oscila entre 5 y 40 por segundo.
La transmisión de cambios en la intensidad lumínica: la respuesta «encendido-apagado», la excitación de muchas células ganglionares depende específicamente de los cambios en la intensidad de la luz. Los impulsos son rápidos durante una fracción de segundo cuando la luz se enciende por primera vez, pero estos impulsos disminuyen rápidamente en la siguiente fracción. El trazado inferior corresponde a una célula ganglionar que ocupa una posición lateral al punto de luz; en este caso, presenta un acusado descenso de su actividad cuando se enciende debido a la inhibición lateral. Después, al apagarse, suceden los efectos opuestos. Por tanto, estos registros se denominan respuestas «encendido-apagado» y «apagado-encendido». Las direcciones contrarias de estas respuestas a la luz están ocasionadas por las células bipolares despolarizantes e hiperpolarizantes, respectivamente, y el origen de la naturaleza transitoria en sus reacciones puede deberse en parte a las células amacrinas, muchas de las cuales ofrecen respuestas pasajeras semejantes.
