Un principio fundamental de la función circulatoria es que la mayoría de los tejidos tienen la capacidad de controlar su propio flujo sanguíneo local en proporción a sus necesidades metabólicas concretas.
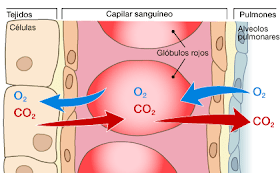
Aporte de oxígeno a los tejidos.
Aporte de otros nutrientes, como glucosa, aminoácidos y ácidos grasos.
Eliminación de dióxido de carbono de los tejidos.
Eliminación de iones hidrógeno de los tejidos.
Mantenimiento de las concentraciones adecuadas de iones en los tejidos.
Transporte de varias hormonas y otras sustancias a los distintos tejidos.
El flujo sanguíneo de la piel determina la pérdida de calor corporal y, de esta forma, se controla la temperatura.
MECANISMOS DE CONTROL DEL FLUJO SANGUÍNEO
El control del flujo sanguíneo local se puede dividir en dos fases: 1) control a corto plazo, y 2) control a largo plazo. El control a corto plazo se consigue con cambios rápidos de la vasodilatación o vasoconstricción local de las arteriolas, metaarteriolas y esfínteres precapilares, que se producen en segundos o minutos para proporcionar con gran rapidez el mantenimiento del flujo sanguíneo tisular local apropiado. El control a largo plazo significa cambios controlados lentos del flujo en un período de días, semanas o incluso meses. En general, estos cambios a largo plazo proporcionan un control aún mejor del flujo en proporción a las necesidades de los tejidos. Estos cambios se producen como consecuencia del incremento o descenso del tamaño físico y del número de vasos sanguíneos que nutren los tejidos.
Uno de los nutrientes metabólicos más necesarios de los tejidos es el oxígeno. El flujo sanguíneo tisular aumenta mucho siempre que disminuye la disponibilidad de oxígeno en los tejidos, por ejemplo: 1) a una gran altitud, en la cima de una montaña alta; 2) en caso de neumonía; 3) en el
Envenenamiento por monóxido de carbono (que deteriora la capacidad de la hemoglobina de transportar el oxígeno), o 4) en el envenenamiento por cianuro.
Teoría vasodilatadora de la regulación a corto plazo del flujo sanguíneo local: posible papel especial de la adenosina.
Teoría de la demanda de oxígeno para el control del flujo sanguíneo local.
Aunque la teoría vasodilatadora es la más aceptada, hay varios hechos críticos que fundamentan otra teoría más expuesta por otros fisiólogos, que podríamos denominar la teoría de la demanda de oxígeno o, más exactamente, la teoría de la demanda de nutrientes (porque están implicados otros nutrientes, además del oxígeno). El oxígeno es uno de los nutrientes metabólicos necesarios para provocar la contracción muscular (con la necesidad también de otros nutrientes). Por tanto, es razonable creer que los vasos sanguíneos simplemente se relajarían en ausencia de una cantidad adecuada de oxígeno, dilatándose de forma natural. Además, el aumento de la utilización de oxígeno en los tejidos como consecuencia del aumento del metabolismo podría, en teoría, disminuir la disponibilidad de oxígeno hacia las fibras musculares lisas de los vasos sanguíneos locales, lo cual también provocaría la vasodilatación local.
La «hiperemia reactiva»
Cuando la sangre que irriga un tejido se bloquea durante unos segundos o durante 1 h o más, y después se desbloquea, el flujo sanguíneo que atraviesa el tejido aumenta inmediatamente hasta 4-7 veces con
Respecto a lo normal; este aumento del flujo continuará durante varios segundos, si el bloqueo ha durado solo unos segundos, pero a veces continuará muchas horas, si el flujo sanguíneo ha estado interrumpido durante 1 h o más. Este fenómeno se conoce como hiperemia reactiva.
La «hiperemia activa»
Cuando cualquier tejido se vuelve muy activo, como un músculo que hace ejercicio, una glándula gastrointestinal durante el período hipersecretor o incluso en el cerebro durante el aumento de la actividad mental, la velocidad del flujo sanguíneo aumenta a través del tejido.
El incremento del metabolismo local hace que las células devoren rápidamente los nutrientes del líquido tisular y también que liberen grandes cantidades de sustancias vasodilatadoras. El resultado es que se dilatan los vasos sanguíneos locales y aumenta el flujo sanguíneo local. De esta forma, el tejido activo recibe los nutrientes adicionales necesarios para mantener este nuevo nivel de funcionamiento.
Autorregulación del flujo sanguíneo durante los cambios en la presión arterial: mecanismos «metabólicos» y «miógenos».
La teoría metabólica se puede entender como: cuando la presión
Arterial es demasiado elevada, el exceso de líquido proporciona demasiado oxígeno y demasiados nutrientes de otro tipo hacia los tejidos y «lava» los vasodilatadores liberados por los tejidos. Estos nutrientes (en especial, el oxígeno), junto con el descenso en los niveles tisulares de vasodilatadores,
Provocan entonces la constricción de los vasos sanguíneos y el retorno del flujo casi a la normalidad, a pesar de que aumente la presión.

Teoría miógena habría otro mecanismo no relacionado con el metabolismo tisular que explicaría el fenómeno de la autorregulación. Esta teoría se basa en la observación de que el estiramiento brusco de los vasos sanguíneos pequeños provoca la contracción del músculo liso de la pared vascular durante unos segundos. Por tanto, se ha propuesto que cuando una presión arterial elevada estira el vaso se provoca, a su vez, una constricción vascular reactiva que reduce el flujo sanguíneo casi a la normalidad.
Regulación a largo plazo del flujo sanguíneo.
La mayoría de los mecanismos de regulación del flujo sanguíneo local que hemos comentado actúan en pocos segundos o minutos después del cambio de la situación tisular local. A pesar de ello, el flujo sanguíneo se ajusta solo en las tres cuartas partes de las necesidades adicionales de los tejidos, incluso después de la activación completa de esos mecanismos agudos. La regulación a largo plazo del flujo sanguíneo es especialmente importante cuando cambian las demandas metabólicas del tejido a largo plazo. Es decir, si un tejido esta crónicamente hiperactivo y, por tanto requiere u aumento crónico de las cantidades de oxigeno y otros nutrientes, por lo que en alguna semanas aumentan tanto el numero como el tamaño de las arteriolas y los vasos capilares para cubrir las necesidades del tejido, a menos que el aparato circulatorio se vuelva patológico o sea demasiado viejo para responder.
Las arterias y arteriolas muy pequeñas de cada tejido mediante la deficiencia de sustancias vasodilatadoras o de oxigeno en las células tisulares solo pueden llegara esos vasos, no a las arterias intermedias y mayores del territorio proximal. Aun así, cuando aumenta el flujo sanguíneo a través de una porción microvascular de la circulación, secundariamente se activa otro mecanismo que dilata también las arterias mayores sintetizan varias sustancias que liberadas afectan el grado de relajación o contracción de la pared arterial.
La noradrenalina es una hormona vasoconstrictora especialmente potente; la adrenalina es menos potente y en algunos tejidos provoca incluso una vasodilatación leve.
Cuando se estimula el sistema nervioso simpático en el cuerpo durante el estrés o el ejercicio, las terminaciones nerviosas simpáticas de cada tejido liberan noradrenalina, que excita al corazón y contrae las venas y las arteriolas. Además, los nervios simpáticos de la médula suprarrenal provocan la secreción de noradrenalina y adrenalina en la sangre. Estas hormonas circulan entonces por todo el cuerpo y provocan casi los mismos efectos en la circulación que la estimulación simpática directa, con
Lo que se consigue un sistema de control doble: 1) estimulación nerviosa directa, y 2) efectos indirectos de la noradrenalina y/o de la adrenalina en la sangre circulante.
Vasopresina
La vasopresina, que también se conoce como hormona antidiurética, es aún más potente que la angiotensina II como vasoconstrictora, por lo que se convierte en una de las sustancias constrictoras más potentes del organismo. Se forma en las células nerviosas del hipotálamo, pero después es transportada distalmente a través de los axones nerviosos hacia la neurohipófisis, donde es finalmente segregada a la sangre.
Bradicidina
Hay un grupo de sustancias denominadas cininas que provocan una vasodilatación potente cuando se forman en la sangre y en los líquidos tisulares de algunos órganos. Las cininas son pequeños polipéptidos que se escinden por enzimas proteolíticas a partir de α2 globulinas del plasma o los líquidos tisulares. Una enzima proteolítica de particular importancia para tal fin es la calicreína, que se encuentra en la sangre y los líquidos tisulares en una forma inactiva. Esta calicreína inactiva se activa por la maceración de la sangre, por la inflamación tisular o por otros efectos químicos o físicos similares. A medida que se va activando la calicreína actúa inmediatamente sobre la α2 globulina para liberar una cinina llamada calidina, que después se convierte en bradicinina gracias a las enzimas tisulares.
Histamina
La histamina se libera esencialmente en todos los tejidos del organismo cuando sufren daños o se inflaman, o cuando se sufre una reacción alérgica. La mayor parte de la histamina deriva de los mastocitos en los tejidos dañados y de los basófilos en sangre. La histamina tiene un efecto vasodilatador potente sobre las arteriolas y, como la bradicinina, puede
Aumentar en gran medida la porosidad capilar permitiendo la pérdida tanto de líquidos como de proteínas plasmáticas hacia los tejidos
