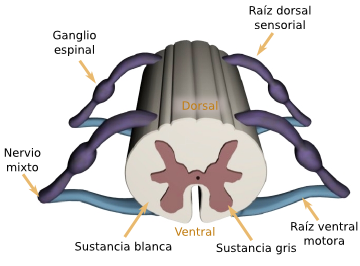
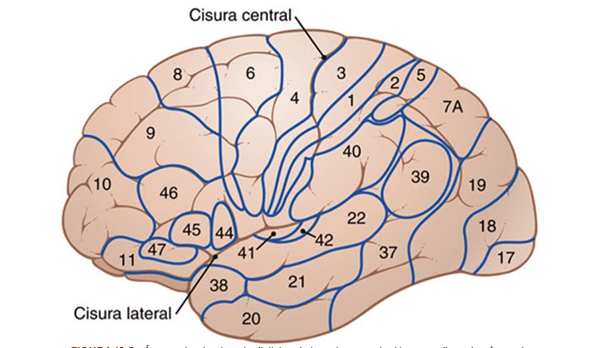
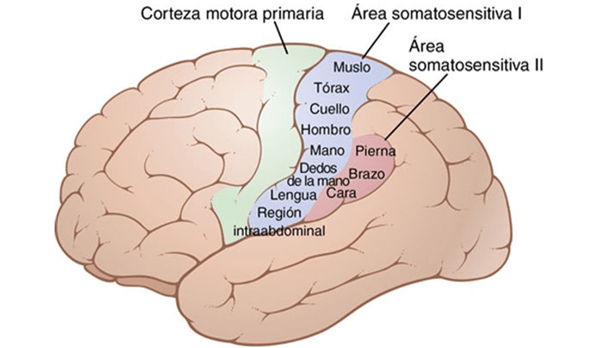
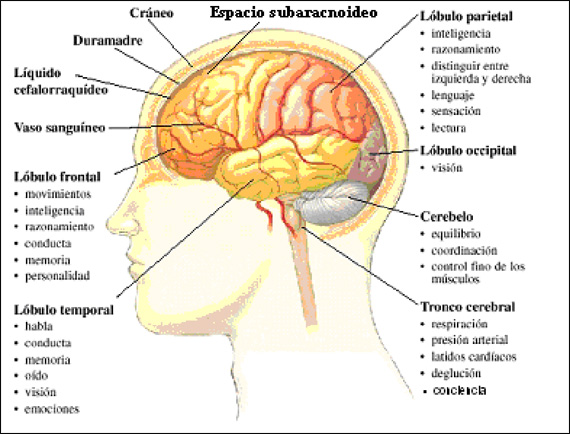
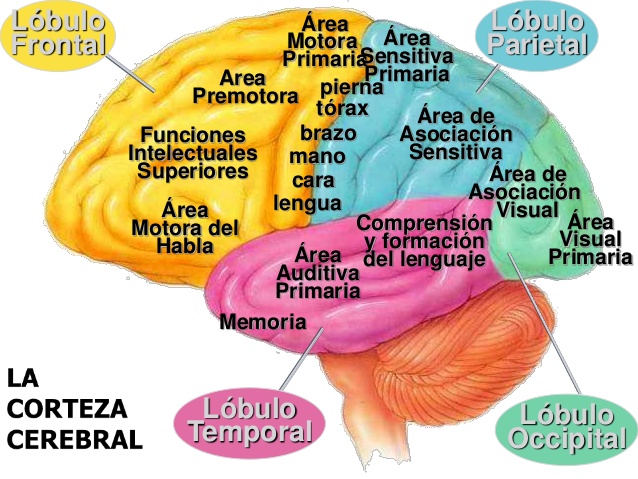
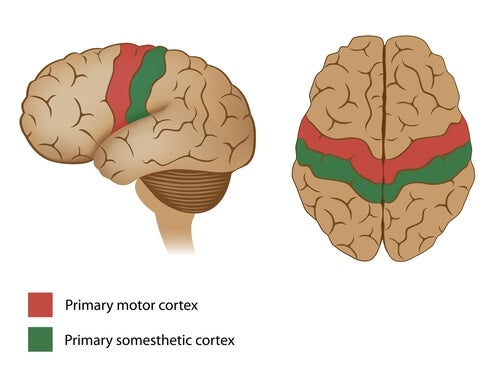
La corteza motora primaria ocupa la primera circunvolución de los lóbulos frontales por delante del surco central o cisura de Rolando. Comienza desde su zona más lateral situada en el surco lateral o cisura de Silvio, se extiende hacia arriba hasta la porción más superior del cerebro y a continuación desciende por la profundidad de la cisura longitudinal.
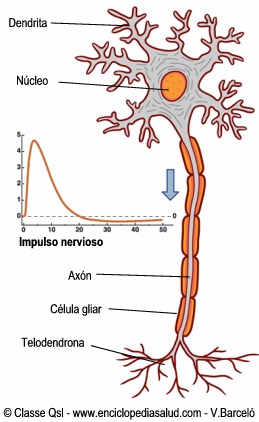
La principal estructura encargada del movimiento, se trata del área que va a generar y enviar la orden de movimiento a todos los músculos voluntarios del organismo. Es la parte del cerebro que envía la orden a los músculos para que se contraigan o tensen, generando el movimiento.
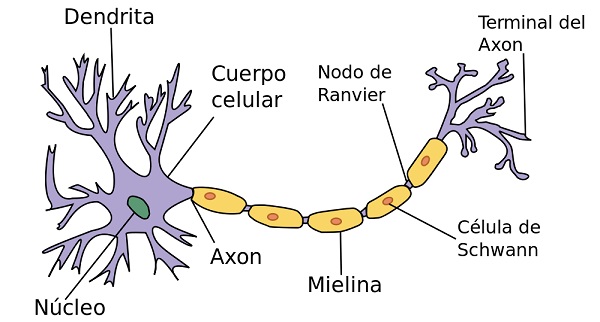
La corteza motora primaria no trabaja sola, sino que precisa de la información proveniente de las siguientes zonas para poder planificar y elaborar los movimientos. En la corteza motora primaria se encuentran, entre otras, las células de Beltz. Estas células de gran longitud, que irán a través de la médula espinal para hacer sinapsis con otras motoneuronas.
El área premotora queda a una distancia de 1 a 3 cm por delante de la corteza motora primaria. Se extiende hacia abajo en dirección al surco lateral y hacia arriba en dirección a la cisura longitudinal, donde limita con el área motora suplementaria, que cumple unas funciones análogas a las del área premotora. La organización topográfica de la corteza premotora es a grandes rasgos la misma que la de la corteza motora primaria, con las zonas para la boca y la cara en una situación más lateral; a medida que se asciende, aparecen las áreas para las manos, los brazos, el tronco y las piernas.
Las señales nerviosas generadas en el área premotora dan lugar a «patrones» de movimiento mucho más complejos que los patrones puntuales originados en la corteza motora primaria
El área motora suplementaria posee otra organización topográfica para controlar la función motora. Sobre todo ocupa la cisura longitudinal, pero se extiende unos pocos centímetros por la corteza frontal superior. Las contracciones suscitadas al estimular esta zona suelen ser bilaterales en vez de unilaterales.
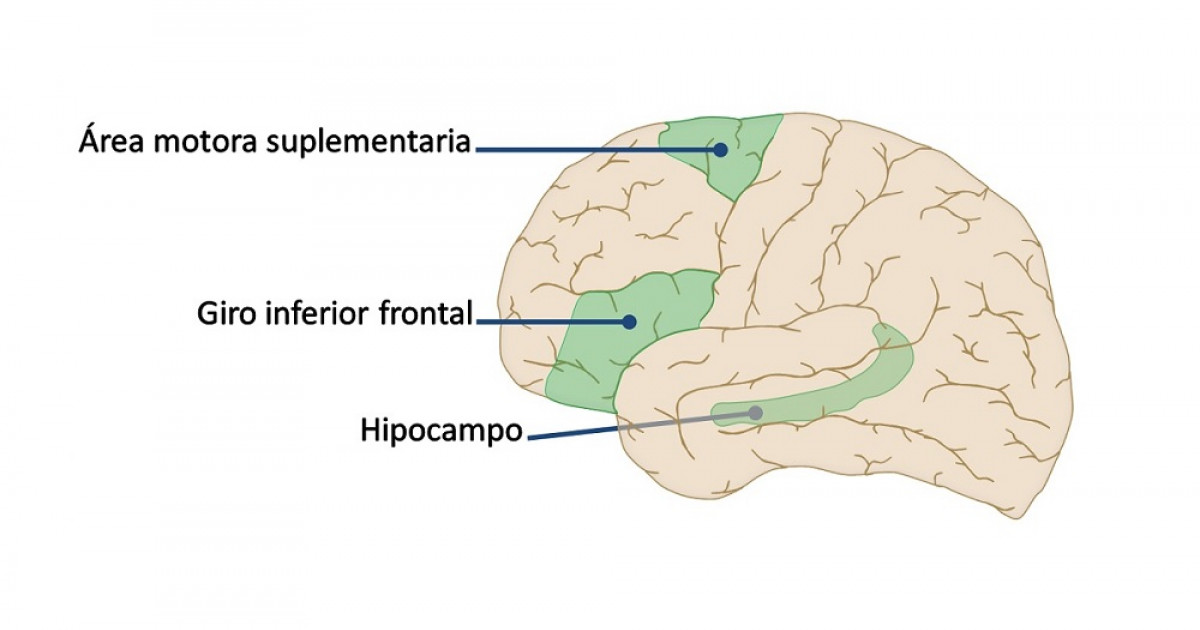
Este área funciona en consonancia con el área premotora para aportar los movimientos posturales de todo el cuerpo, los movimientos de fijación de los diversos segmentos corporales, los movimientos posturales de la cabeza y de los ojos.

Área de Broca un área premotora designada con la expresión «formación de las palabras» que se halla justo delante de la corteza motora primaria e inmediatamente por encima del surco lateral. Su lesión no impide que una persona vocalice, pero hace imposible que emita palabras completas en vez de sonidos descoordinados o algún término sencillo esporádico.
Los movimientos oculares voluntarios. En el área premotora justo por encima del área de Broca existe un punto encargado de controlar los movimientos voluntarios de los ojos. Su lesión impide a una persona dirigirlos de forma voluntaria hacia los diversos objetos
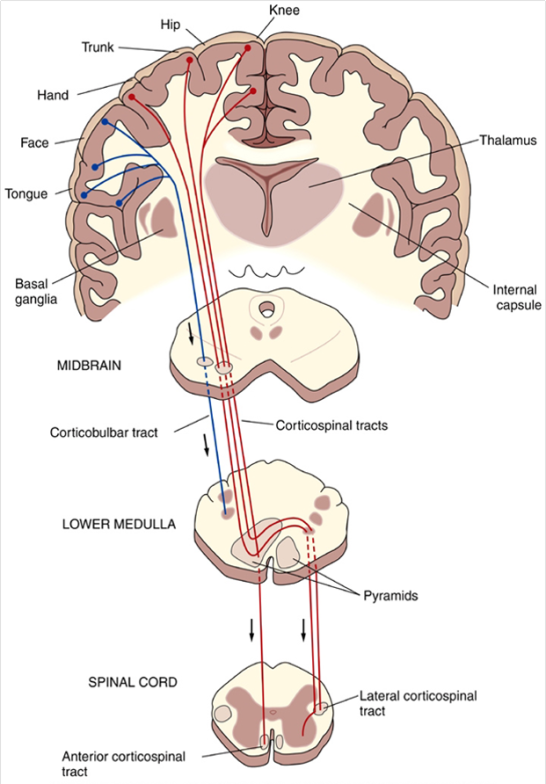
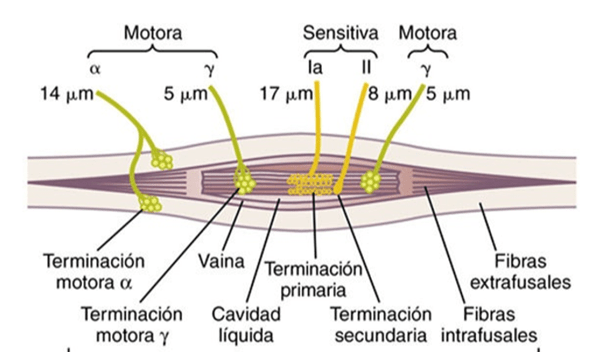
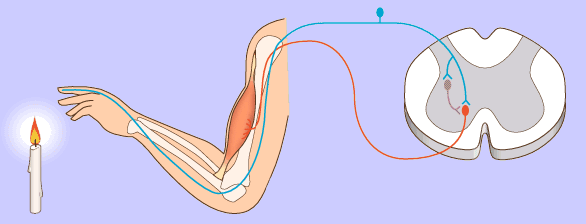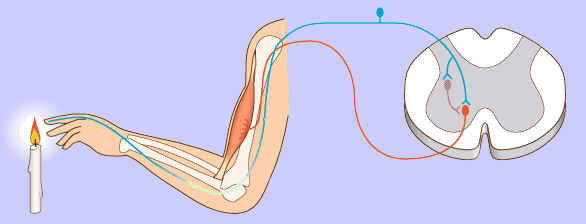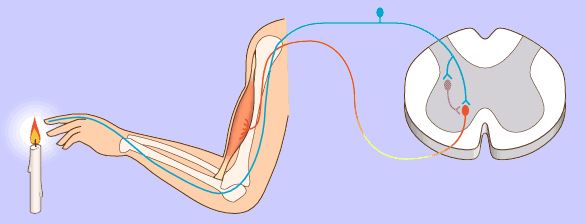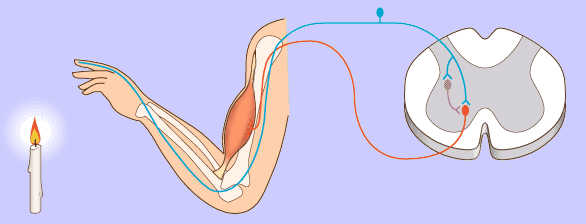
Las señales motoras se transmiten directamente desde la corteza hasta la médula espinal a través del fascículo corticoespinal e indirectamente por múltiples vías accesorias en las que intervienen los ganglios basales, el cerebelo y diversos núcleos del tronco del encéfalo. Las vías directas están más dedicadas a los movimientos detallados y bien diferenciados, especialmente en los segmentos distales de las extremidades, sobre todo en las manos y los dedos.
Las neuronas pertenecientes a cada columna operan como un sistema de procesamiento integrado, manejan información procedente de múltiples fuentes para determinar la respuesta emitida por la columna. cada columna puede funcionar como un sistema amplificador para estimular una gran cantidad de fibras piramidales dirigidas al mismo músculo o a los músculos sinérgicos en un momento dado.
Esta capacidad es importante, porque la activación de una sola célula piramidal rara
vez es capaz de excitar un músculo. Hace falta la excitación de 50 a 100 simultáneamente o en una rápida sucesión para lograr la contracción muscular definitiva.
El tronco del encéfalo es dueño de sí mismo, porque se encarga de muchas funciones de control especiales, como:
1. Control de la respiración
2. Control del aparato cardiovascular
3. Control parcial del funcionamiento digestivo
4. Control de muchos movimientos estereotipados del cuerpo
5. Control del equilibrio
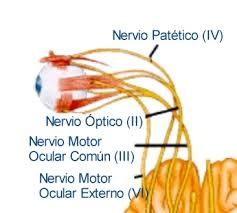
6. Control de los movimientos oculares
El tronco del encéfalo sirve como estación de relevo para las «señales de mando» procedentes de los centros nerviosos superiores.

Los núcleos reticulares pontinos transmiten señales excitadoras en sentido descendente hacia la médula a través del fascículo reticuloespinal pontino situado en la columna anterior de esta estructura. Las fibras de esta vía terminan sobre las motoneuronas anteriores mediales que activan a los músculos axiales del cuerpo, los que lo sostienen en contra de la gravedad y que corresponden a los músculos de la columna vertebral y los extensores de las extremidades.
Los núcleos reticulares bulbares transmiten señales inhibidoras hacia las mismas motoneuronas anteriores antigravitatorias a través de una vía diferente, el fascículo reticuloespinal bulbar, situado en la columna lateral de la médula. Los núcleos reticulares bulbares reciben potentes colaterales aferentes desde:
1) el fascículo corticoespinal
2) el fascículo rubroespinal
3) otras vías motoras.
Estos fascículos y vías normalmente activan este sistema reticular bulbar de carácter inhibidor para compensar las señales excitadoras del sistema reticular pontino, por lo que, en condiciones normales, los músculos del cuerpo no presentan una tirantez anormal.

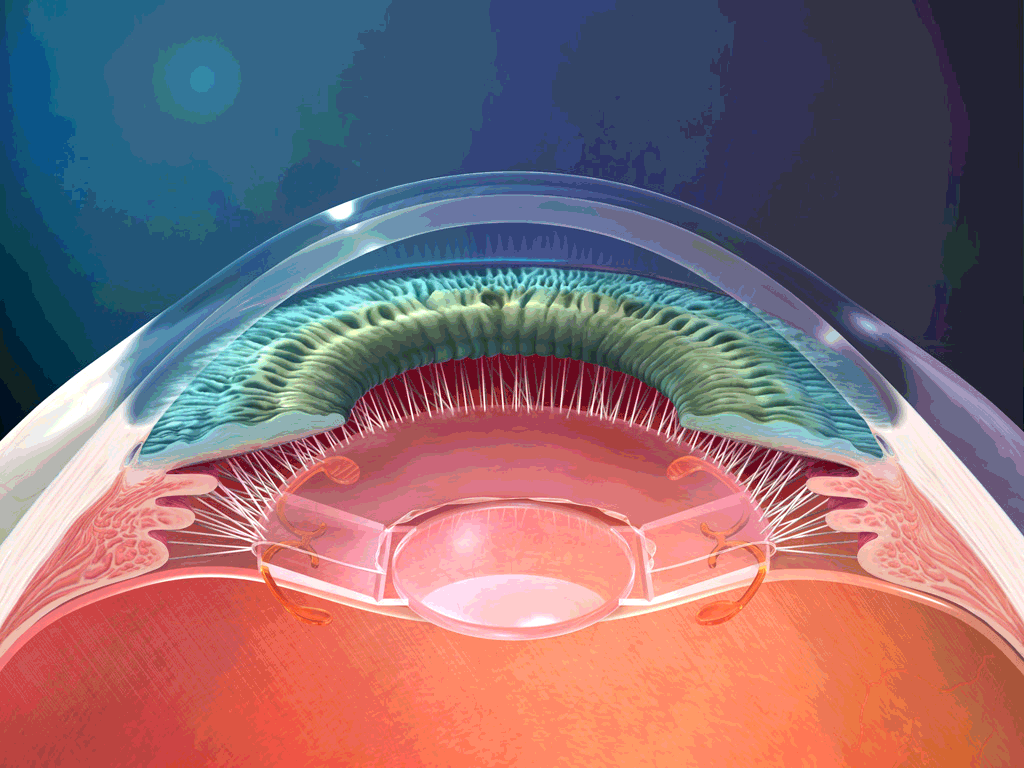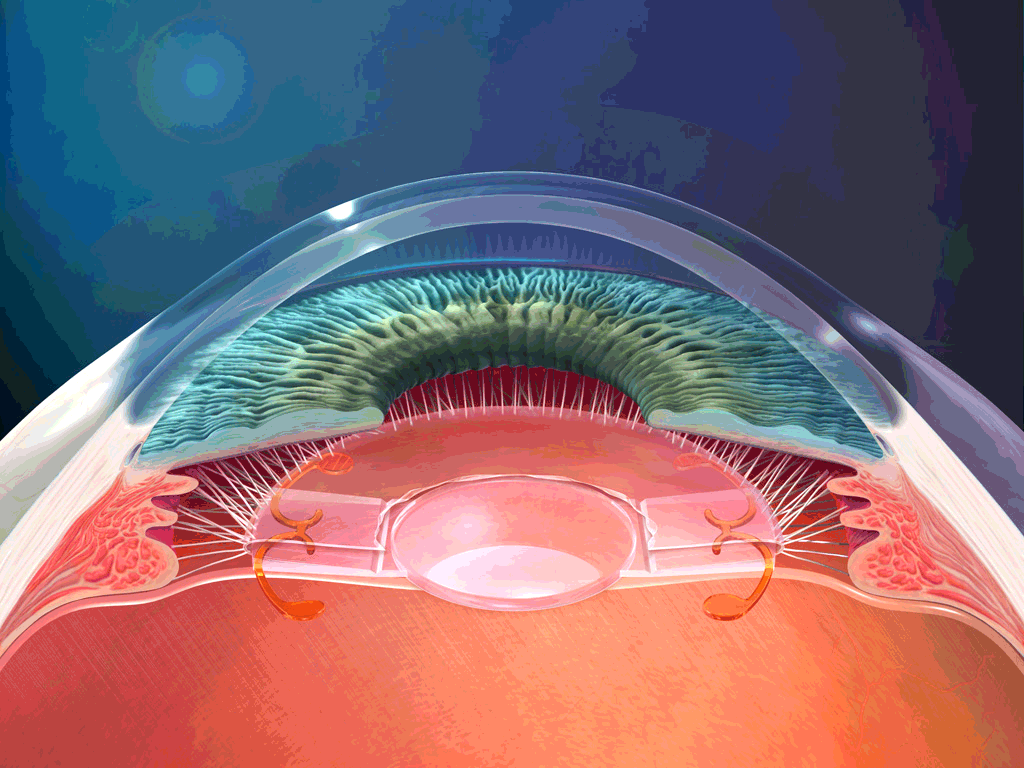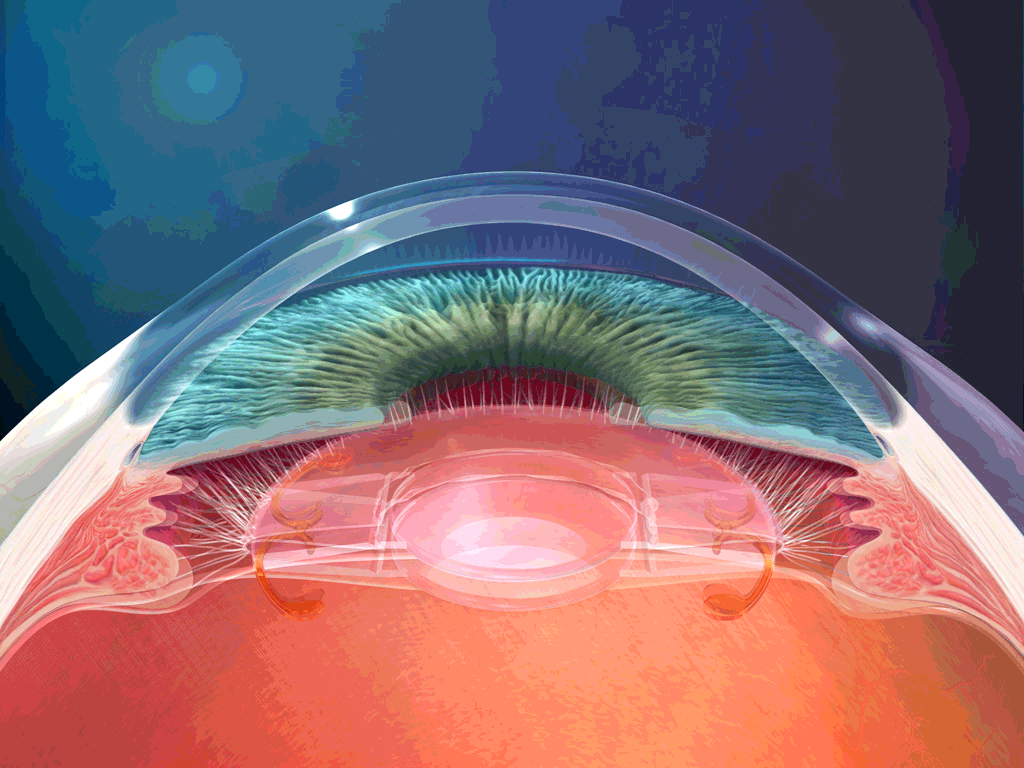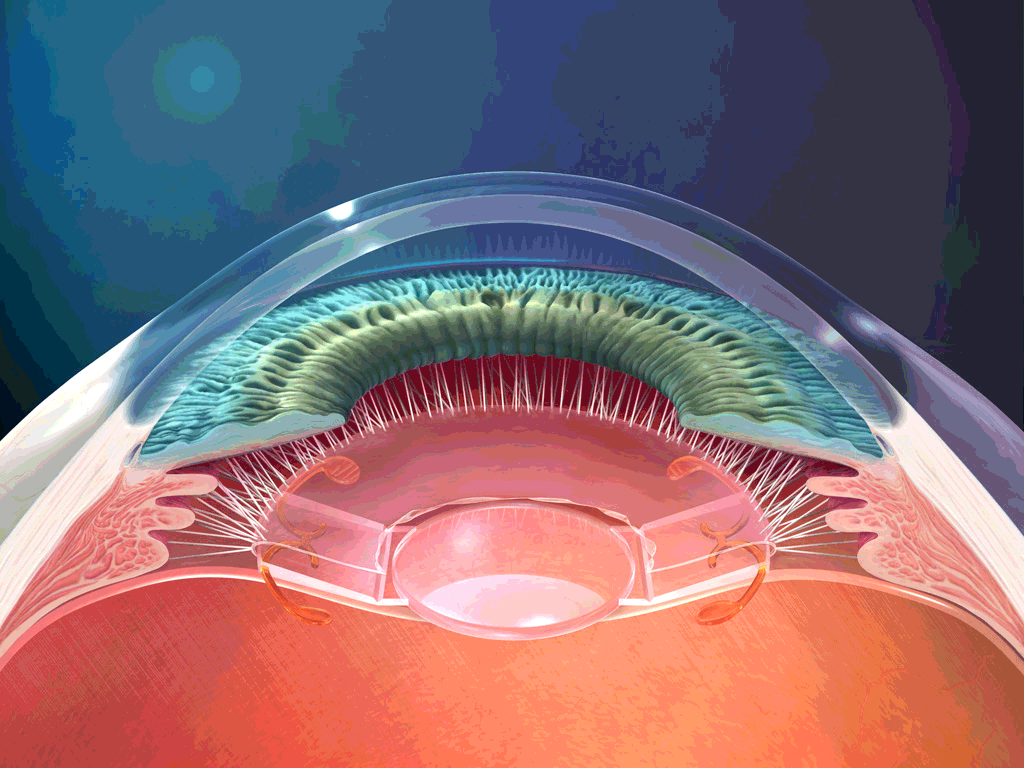
El aparato vestibular, es el órgano sensitivo encargado de detectar la sensación del equilibrio. Se encuentra encerrado en un sistema de tubos y cavidades óseas situado en la porción petrosa del hueso temporal, llamado laberinto óseo. Dentro de este sistema están los tubos y cavidades membranosas denominados laberinto membranoso. El laberinto membranoso es el componente funcional del aparato vestibular.
La mácula del utrículo queda básicamente en el plano horizontal de la superficie inferior del utrículo y cumple una función importante para determinar la orientación de la cabeza cuando se encuentra en posición vertical. En líneas generales la mácula del sáculo está situada en un plano vertical e informa de la orientación de la cabeza cuando la persona está tumbada.
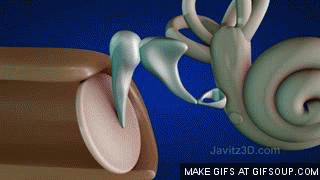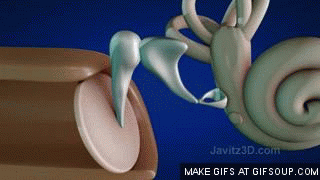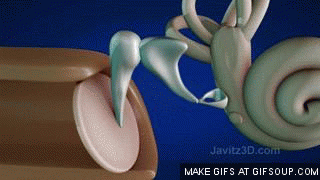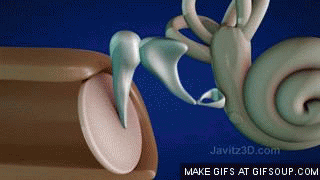
La mácula se encuentra cubierta por una capa gelatinosa en la que están enterrados muchos pequeños cristales de carbonato cálcico llamados otolitos o estatoconias.
Los conductos semicirculares anterior, posterior y lateral (horizontal) mantienen una disposición perpendicular entre sí de manera que representan los tres planos del espacio. Cuando la cabeza se inclina hacia delante unos 30°, los conductos semicirculares laterales quedan aproximadamente horizontales con respecto a la superficie del suelo; los anteriores están en un plano vertical que se proyecta hacia delante y 45° hacia fuera, mientras que los posteriores están en planos verticales que se proyectan hacia atrás y 45° hacia fuera.
El sáculo y utrículo, son llamados laberintos estáticos, ya que su función está relacionada con la detección de la posición de la cabeza respecto a la gravedad, y no al movimiento voluntario. Es por esto que pequeños cambios en la posición de la cabeza distorsionan y estimulan las células ciliadas.