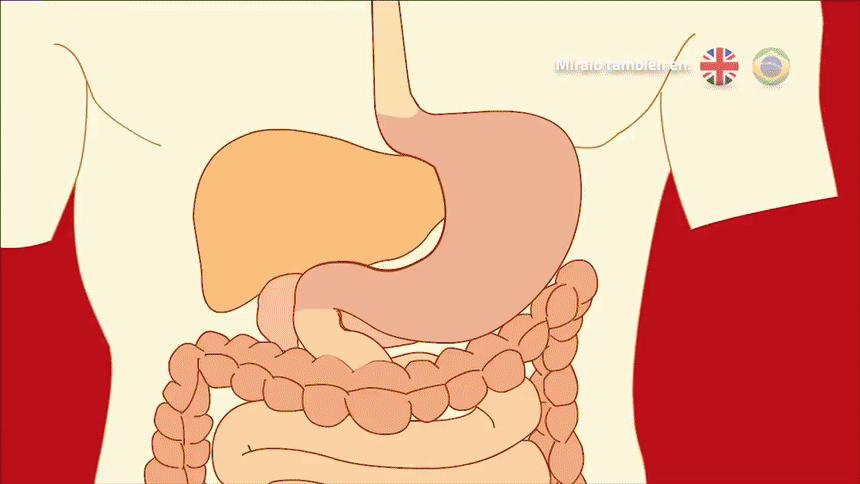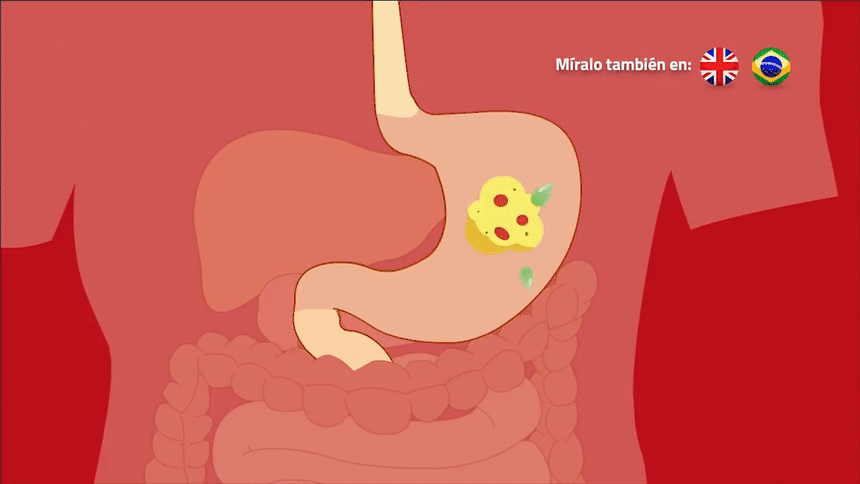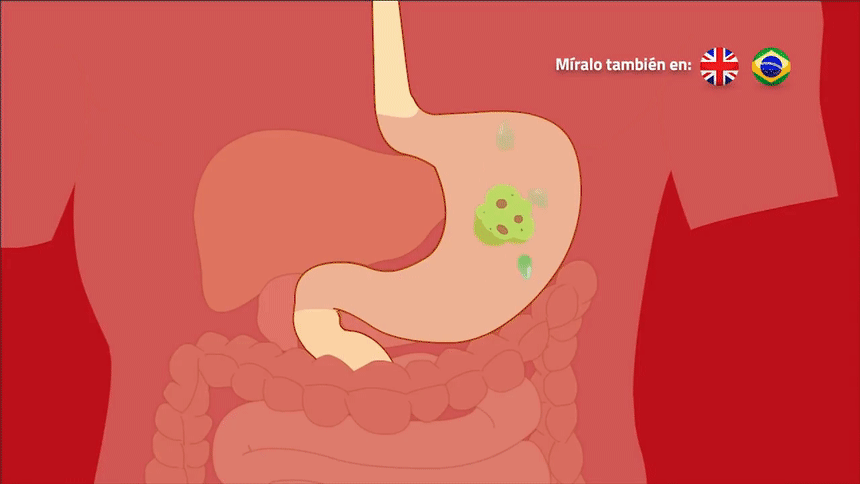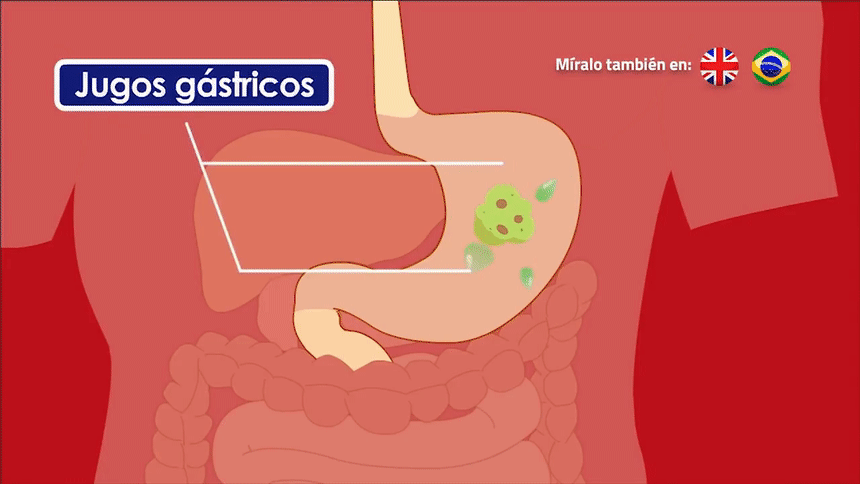
músculo liso, que está formado por fibras mucho menores, habitualmente de 1 a 5 μm de diámetro y de solo 20 a 500 μm de longitud. Por el contrario, las fibras musculares esqueléticas tienen un diámetro hasta 30 veces mayor y una longitud cientos de veces mayor. Muchos de los mismos principios de la contracción se aplican al músculo liso y al músculo esquelético. Lo que es más importante, esencialmente las mismas fuerzas de atracción entre los filamentos de miosina y actina producen la contracción en el músculo liso y en el músculo esquelético, pero la disposición física interna de las fibras musculares lisas es diferente.
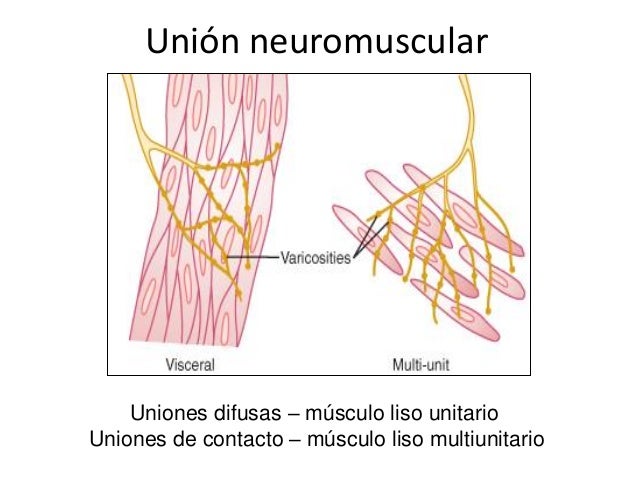
Músculo liso multiunitario Este tipo de músculo liso está formado por fibras musculares lisas separadas y discretas. Cada una de las fibras actúa independientemente de las demás y con frecuencia está inervada por una única terminación nerviosa, como ocurre en las fibras musculares esqueléticas.
Además, la superficie externa de estas fibras, como en el caso de las fibras musculares esqueléticas, está cubierta por una capa delgada de sustancia similar a una membrana basal, una mezcla de colágeno fino y glucoproteínas que aísla las fibras separadas entre sí. Las características fundamentales de las fibras musculares lisas multiunitarias son que cada una de las fibras se puede contraer independientemente de las demás, y que su control se ejerce principalmente por señales nerviosas. Por el contrario, una parte importante del control del músculo liso unitario es ejercida por estímulos no nerviosos.
Músculo liso unitario Este tipo se denomina músculo liso sincitial o músculo liso visceral. El término «unitario» es confuso porque no se refiere a fibras musculares únicas. Por el contrario, se refiere a una masa de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad. Las fibras habitualmente están dispuestas en láminas o fascículos, y sus membranas celulares están adheridas entre sí en múltiples puntos, de modo que la fuerza que se genera en una fibra muscular se puede transmitir a la siguiente.
En lugar de la troponina, las células musculares lisas contienen una gran cantidad de otra proteína reguladora denominada calmodulina. Aunque esta proteína es similar a la troponina, inicia la contracción de una manera diferente. La calmodulina inicia la contracción al activar los puentes cruzados de miosina. Esta activación y la posterior contracción se producen según la siguiente secuencia:
1. La concentración de calcio en el líquido citosólico del músculo liso se incrementa como consecuencia de la entrada de calcio desde el líquido extracelular a través de los canales de calcio y/o la liberación de calcio desde el retículo sarcoplásmico.
2. Los iones calcio se unen a la calmodulina de forma reversible.
3. El complejo calmodulina-calcio se une después a la miosina cinasa de cadena ligera, que es una enzima fosforiladora, y la activa.
4. Una de las cadenas ligeras de cada una de las cabezas de miosina, denominada cabeza reguladora, se fosforila en respuesta a esta miosina cinasa. Cuando esta cadena no está fosforilada no se produce el ciclo de unión-separación de la cabeza de miosina con el filamento de actina. Sin embargo, cuando la cadena reguladora está fosforilada la cabeza tiene la capacidad de unirse repetitivamente al filamento de actina y de avanzar a través de todo el proceso de ciclado de «tirones» intermitentes, al igual que ocurre en el músculo esquelético, produciendo de esta manera la contracción muscular.
La miosina fosfatasa es importante en la interrupción de la contracción La relajación del músculo liso tiene lugar cuando los canales de calcio se cierran y la bomba de calcio transporta iones calcio fuera del líquido citosólico de la célula. Cuando la concentración de iones calcio disminuye por debajo de un nivel crítico, los procesos que se acaban de señalar se invierten automáticamente, excepto la fosforilación de la cabeza de miosina. La inversión de esta reacción precisa otra enzima, la miosina fosfatasa, que está localizada en el citosol de la célula muscular lisa y que escinde el fosfato de la cadena ligera reguladora. Después se interrumpe el ciclo y finaliza la contracción.
Potenciales de membrana en el músculo liso
El voltaje cuantitativo del potencial de membrana del músculo liso depende de la situación momentánea del músculo. En el estado de reposo normal el potencial intracelular es de aproximadamente –50 a –60 mV , alrededor de 30 mV menos negativo que en el músculo esquelético.
Potenciales de acción en el músculo liso unitario
Los potenciales de acción se producen en el músculo liso unitario (como el músculo visceral) de la misma forma que en el músculo esquelético. Normalmente no se producen en la mayoría de los tipos multiunitarios de músculo liso, como se analiza en una sección posterior. Los potenciales de acción del músculo liso visceral se producen en una de dos formas: 1) potenciales en espiga, o 2) potenciales de acción con meseta.
Potenciales en espiga
Los potenciales de acción en espiga típicos, como los que se ven en el músculo esquelético, aparecen en la mayoría de los tipos de músculo liso unitario. La duración de este tipo de potencial de acción es de 10 a 50 ms, como se ve en la figura 8-7A. Estos potenciales de acción se pueden generar de muchas maneras, por ejemplo mediante estimulación eléctrica, por la acción de hormonas sobre el músculo liso, por la acción de sustancias transmisoras procedentes de las fibras nerviosas.
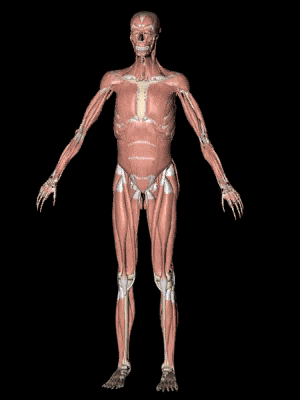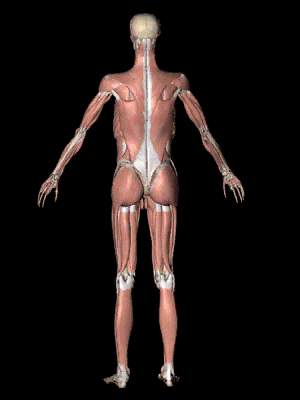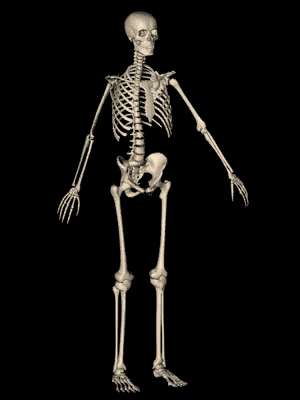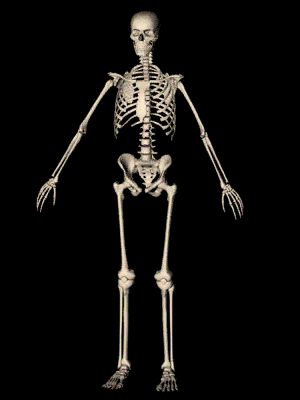
los músculos esqueléticos están formados por numerosas fibras cuyo diámetro varía entre 10 y 80 μm. Cada una de estas fibras está formada por subunidades cada vez más pequeñas.
El sarcolema está formado por una membrana celular verdadera, denominada membrana plasmática, y una cubierta externa formada por una capa delgada de material polisacárido que contiene numerosas fibrillas delgadas de colágeno. En cada uno de los dos extremos de la fibra muscular la capa superficial del sarcolema se fusiona con una fibra tendinosa. Las fibras tendinosas a su vez se agrupan en haces para formar los tendones musculares, que después insertan los músculos en los huesos.
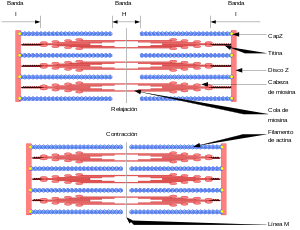
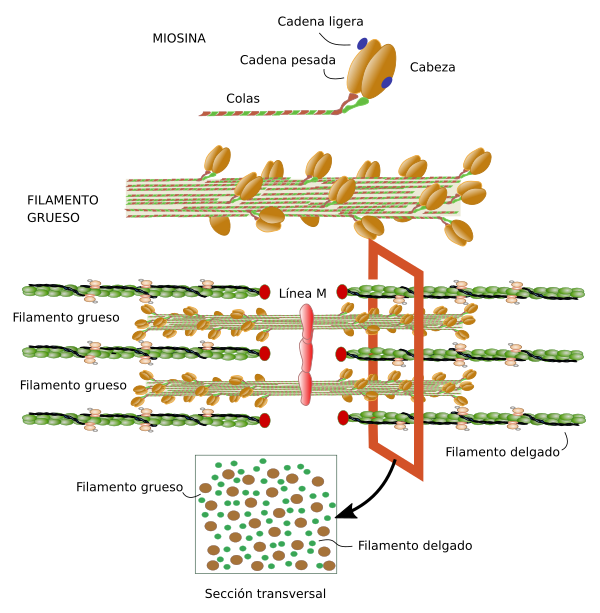
Las miofibrillas están formadas por filamentos de actina y miosina Cada fibra muscular contiene varios cientos a varios miles de miofibrillas, Cada miofibrilla está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina adyacentes entre sí, que son grandes moléculas proteicas polimerizadas responsables de la contracción muscular real.
Mecanismo general de la contracción muscular:
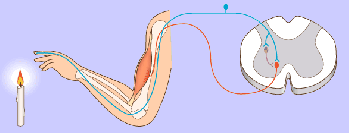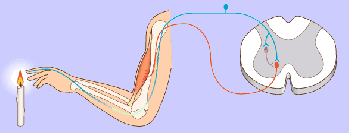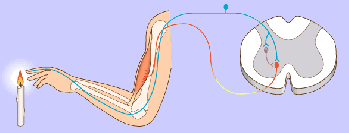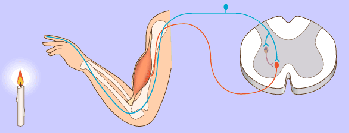
Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares.
2. En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.
3. La acetilcolina actúa en una zona local de la membrana de la fibra muscular para abrir múltiples canales de cationes «activados por acetilcolina» a través de moléculas proteicas que flotan en la membrana.
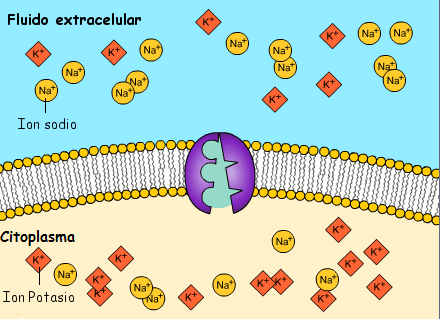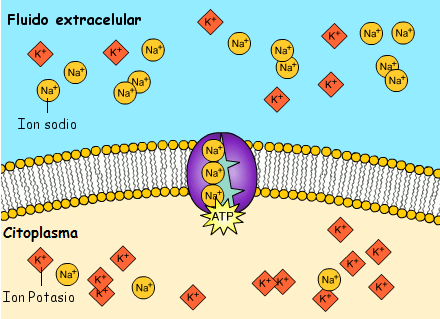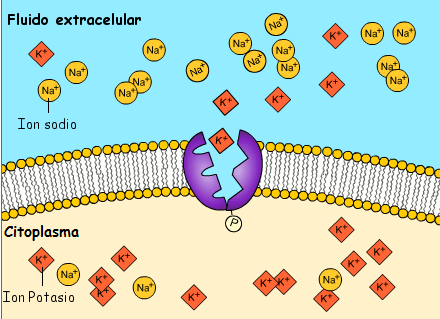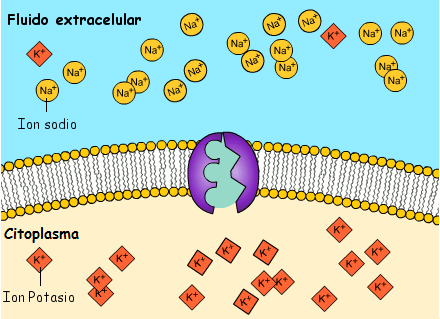
4. La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular. Esta acción provoca una despolarización local que, a su vez, conduce a la apertura de los canales de sodio activados por el voltaje, que inicia un potencial de acción en la membrana.
5. El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas.
6. El potencial de acción despolariza la membrana muscular, y buena parte de la electricidad del potencial de acción fluye a través del centro de la fibra muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de iones calcio que se han almacenado en el interior de este retículo.
7. Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslicen unos sobre otros en sentido longitudinal, lo que constituye el proceso contráctil.
8. Después de una fracción de segundo los iones calcio son bombeados de nuevo hacia el retículo sarcoplásmico por una bomba de Ca++ de la membrana y permanecen almacenados en el retículo hasta que llega un nuevo potencial de acción muscular; esta retirada de los iones calcio desde las miofibrillas hace que cese la contracción muscular.
ATP como fuente de energía para la contracción:
Antes de que comience la contracción, las cabezas de los puentes cruzados se unen al ATP. La actividad ATPasa de la cabeza de miosina escinde inmediatamente el ATP, aunque deja los productos de la escisión, el ADP y el ion fosfato, unidos a la cabeza. En este estado la conformación de la cabeza es tal que se extiende perpendicularmente hacia el filamento de actina, pero todavía no está unida a ella.
2. Cuando el complejo troponina-tropomiosina se une a los iones calcio quedan al descubierto los puntos activos del filamento de actina, y entonces las cabezas de miosina se unen a estos sitios
3. El enlace entre la cabeza del puente cruzado y el punto activo del filamento de actina produce un cambio conformacional de la cabeza, lo que hace que la cabeza se desplace hacia el brazo del puente cruzado, lo que proporciona el golpe activo para tirar del filamento de actina. La energía que activa el golpe activo es la energía que ya se ha almacenado, como un muelle «comprimido» por el cambio conformacional que se había producido previamente en la cabeza cuando se escindió la molécula de ATP.
4. Una vez que se desplaza la cabeza del puente cruzado, se facilita la liberación del ADP y el ion fosfato que previamente estaban unidos a la cabeza. En el punto de liberación del ADP se une una nueva molécula de ATP, lo cual hace que la cabeza se separe de la actina.
5. Después de que la cabeza se haya separado de la actina, se escinde la nueva molécula de ATP para comenzar el ciclo siguiente, dando lugar a un nuevo golpe activo. Es decir, la energía «comprime» la cabeza una vez más a su situación perpendicular, dispuesta para comenzar el nuevo ciclo de golpe activo.
6. Cuando la cabeza comprimida (con su energía almacenada procedente del ATP escindido) se une a un nuevo punto activo del filamento de actina, se estira y una vez más proporciona un nuevo golpe activo. De esta manera el proceso se realiza una y otra vez hasta que los filamentos de actina han desplazado la membrana Z hasta los extremos de los filamentos de miosina o hasta que la carga que se ejerce sobre el músculo se hace demasiado grande como para que se produzca una tracción adicional.
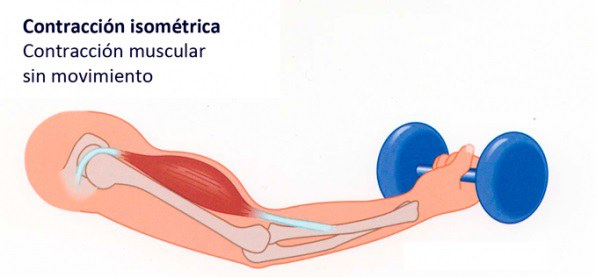
Las contracciones isométricas no acortan el músculo, mientras que las contracciones isotónicas lo acortan a una tensión constante Se dice que la contracción muscular es isométrica cuando el músculo no se acorta durante la contracción e isotónica cuando se acorta, pero la tensión del músculo permanece constante durante toda la contracción.
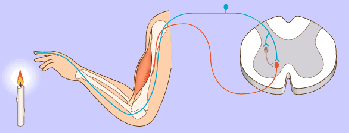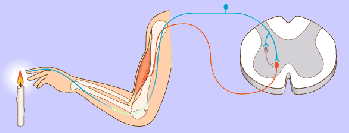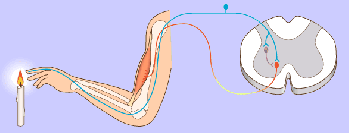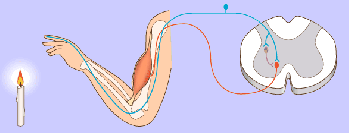
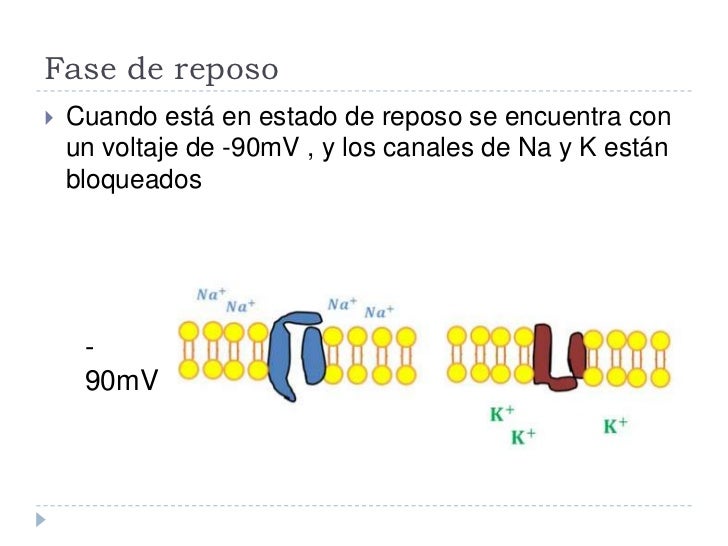
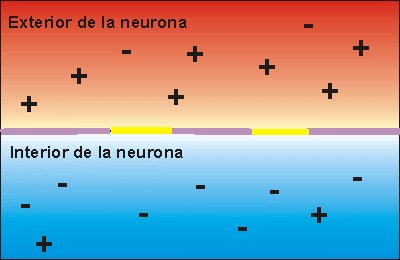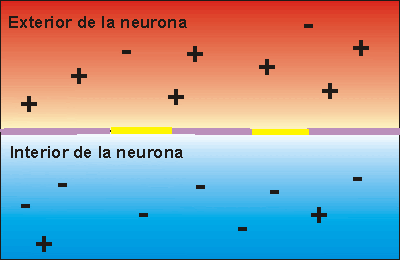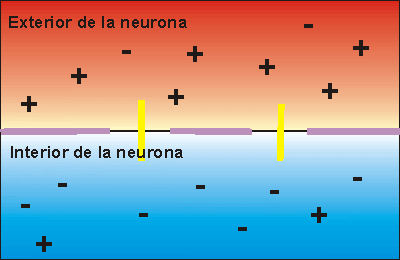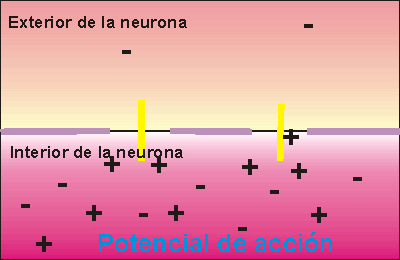
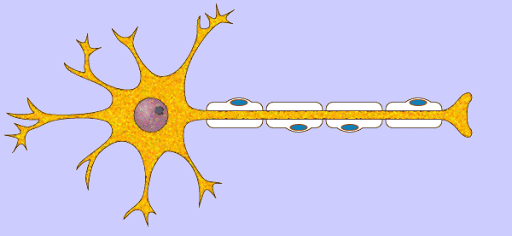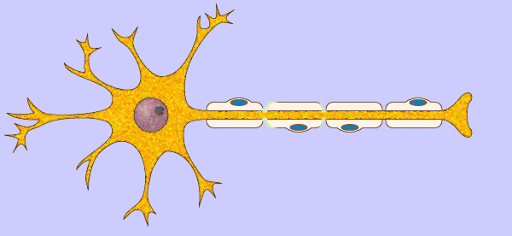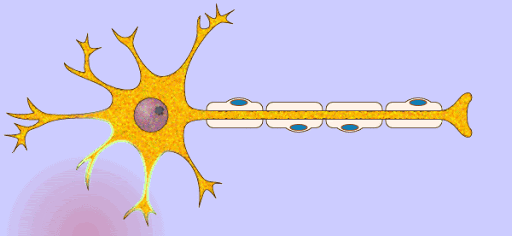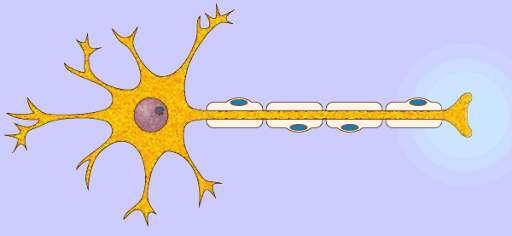
Potencial de membrana en reposo de las neuronas. El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90 mV . Es decir, el potencial en el interior de la fibra es 90 mV más negativo que el potencial del líquido extracelular que está en el exterior de la misma.
Potencial de acción de las neuronas:
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega a su extremo.
fases sucesivas del potencial de acción:
Fase de reposo: La fase de reposo es el potencial de membrana en reposo antes del comienzo del potencial de acción. Se dice que la membrana está «polarizada» durante esta fase debido al potencial de membrana negativo de –90 mV que está presente.
Fase de despolarización:
la membrana se hace súbitamente muy permeable a los iones sodio, lo que permite que un gran número de iones sodio con carga positiva difunda hacia el interior del axón. El estado «polarizado» normal de –90 mV se neutraliza inmediatamente por la entrada de iones sodio cargados positivamente, y el potencial aumenta rápidamente en dirección positiva, un proceso denominado despolarización. En las fibras nerviosas grandes el gran exceso de iones sodio positivos que se mueven hacia el interior hace que el potencial de membrana realmente se «sobreexcite» más allá del nivel cero y que se haga algo positivo.
Fase de repolarización: En un plazo de algunas diezmilésimas de segundo después de que la membrana se haya hecho muy permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los canales de potasio se abren más de lo normal. De esta manera, la rápida difusión de los iones potasio hacia el exterior restablece el potencial de membrana en reposo negativo normal, que se denomina repolarización de la membrana.
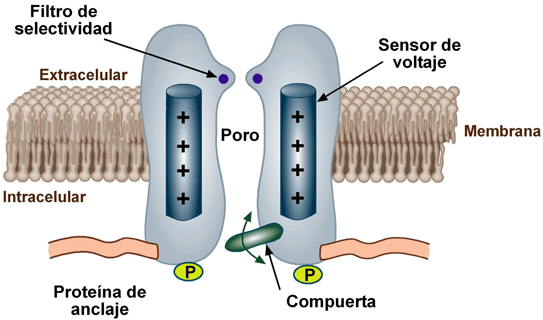
Canales de sodio y potasio activados por el voltaje:
El actor necesario en la producción tanto de la despolarización como de la repolarización de la membrana nerviosa durante el potencial de acción es el canal de sodio activado por el voltaje. Un canal de potasio activado por el voltaje también tiene una función importante en el aumento de la rapidez de la repolarización de la membrana. Estos dos canales activados por el voltaje tienen una función adicional a la de la bomba Na+-K+ y de los canales de fuga de K+.
Principio del todo o nada:
Una vez que se ha originado un potencial de acción en cualquier punto de la membrana de una fibra normal, el proceso de despolarización viaja por toda la membrana si las condiciones son las adecuadas, o no viaja en absoluto si no lo son. Este principio se denomina principio del todo o nada y se aplica a todos los tejidos excitables normales. De manera ocasional, el potencial de acción alcanza un punto de la membrana en el que no genera un voltaje suficiente como para estimular la siguiente zona de la membrana. Cuando se produce esta situación se interrumpe la diseminación de la despolarización. Por tanto, para que se produzca la propagación continuada de un impulso, el cociente del potencial de acción respecto al umbral de excitación debe ser mayor de 1 en todo momento. Este requisito de «mayor de 1» se denomina factor de seguridad para la propagación.
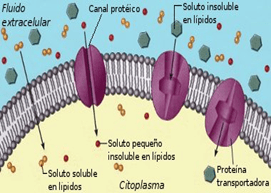
La membrana está formada casi totalmente por una bicapa lipídica, aunque también contiene grandes números de moléculas proteicas insertadas en los lípidos, muchas de las cuales penetran en todo el grosor de la membrana, La bicapa lipídica no es miscible con el líquido extracelular ni con el líquido intracelular.
Las moléculas proteicas de la membrana tienen unas propiedades totalmente diferentes para transportar sustancias. Sus estructuras moleculares interrumpen la continuidad de la bicapa lipídica y constituyen una ruta alternativa a través de la membrana celular. Muchas de estas proteínas penetrantes pueden actuar como proteínas transportadoras. Proteínas diferentes actúan de una manera diferente. Algunas tienen espacios acuosos en todo el trayecto del interior de la molécula y permiten el movimiento libre de agua, así como de iones o moléculas seleccionados; estas proteínas se denominan proteínas de los canales.
El transporte a través de la membrana celular, ya sea directamente a través de la bicapa lipídica o a través de las proteínas, se produce mediante uno de dos procesos básicos: difusión o transporte activo.
Difusión :Todas las moléculas e iones de los líquidos corporales, incluyendo las moléculas de agua y las sustancias disueltas, están en movimiento constante, de modo que cada partícula se mueve de manera completamente independiente.
La difusión a través de la membrana celular se divide en dos subtipos, denominados difusión simple y difusión facilitada.
Difusión simple significa que el movimiento cinético de las moléculas o de los iones se produce a través de una abertura de la membrana o a través de espacios intermoleculares sin ninguna interacción con las proteínas transportadoras de la membrana. La velocidad de difusión viene determinada por la cantidad de sustancia disponible, la velocidad del movimiento cinético y el número y el tamaño de las aberturas de la membrana a través de las cuales se pueden mover las moléculas o los iones
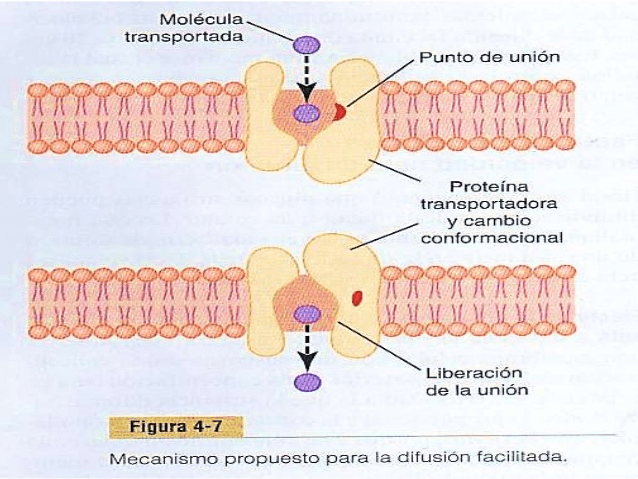
La difusión facilitada precisa la interacción de una proteína transportadora. La proteína transportadora ayuda al paso de las moléculas o de los iones a través de la membrana mediante su unión química con estos y su desplazamiento a través de la membrana de esta manera. Se puede producir difusión simple a través de la membrana celular por dos rutas: 1) a través de los intersticios de la bicapa lipídica si la sustancia que difunde es liposoluble, y 2) a través de canales acuosos que penetran en todo el grosor de la bicapa a través de las grandes proteínas transportadoras.
Un factor importante que determina la rapidez con la que una sustancia difunde a través de la bicapa lipídica es la liposolubilidad.
La difusión facilitada también se denomina difusión mediada por un transportador porque una sustancia que se transporta de esta manera difunde a través de la membrana con la ayuda de una proteína transportadora específica para contribuir al transporte. Es decir, el transportador facilita la difusión de la sustancia hasta el otro lado.
¿Qué limita la velocidad de la difusión facilitada?
La molécula que se va a transportar entra en el poro y queda unida. Después, en una fracción de segundo se produce un cambio conformacional o químico en la proteína transportadora, de modo que el poro ahora se abre en el lado opuesto de la membrana. Como la fuerza de unión del receptor es débil, el movimiento térmico de la molécula unida hace que se separe y que se libere en el lado opuesto de la membrana, La velocidad a la que se pueden transportar moléculas por este mecanismo nunca puede ser mayor que la velocidad a la que la molécula proteica transportadora puede experimentar el cambio en un sentido y en otro entre sus dos estados.
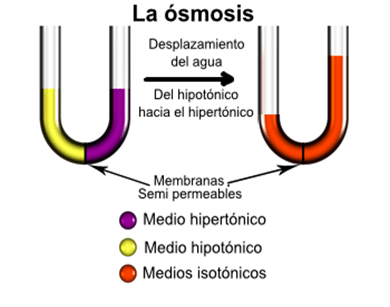
Ósmosis: la sustancia más abundante que difunde a través de la membrana celular es el agua. Cada segundo difunde normalmente una cantidad suficiente de agua en ambas direcciones a través de la membrana del eritrocito igual a aproximadamente 100 veces el volumen de la propia célula, Este proceso de movimiento neto del agua que se debe a la producción de una diferencia de la concentración del agua se denomina ósmosis.
Las moléculas de agua atraviesan la membrana celular con facilidad, mientras que los iones sodio y cloruro pasan solo con dificultad. Por tanto, la solución de cloruro sódico es realmente una mezcla de moléculas de agua difusibles y de iones sodio y cloruro no difusibles, y se dice que la membrana es permeable de manera selectiva al agua, pero mucho menos a los iones sodio y cloruro.
Presión osmótica: La cantidad de presión necesaria para detener la ósmosis se denomina presión osmótica de la solución de cloruro sódico, Esta diferencia de presión a través de la membrana en este punto es igual a la presión osmótica de la solución que contiene el soluto no difusible.
La presión osmótica que ejercen las partículas de una solución, ya sean moléculas o iones, está determinada por el número de partículas por unidad de volumen del líquido, no por la masa de las partículas,
«Osmolalidad»: el osmol, Para expresar la concentración de una solución en función del número de partículas se utiliza la unidad denominada osmol en lugar de los gramos. Un osmol es el peso molecular-gramo de un soluto osmóticamente activo. Relación entre osmolalidad y presión osmótica A la temperatura corporal normal, 37 °C, una concentración de un osmol por litro producirá una presión osmótica de 19.300 mmHg en la solución.
El término «osmolaridad» Osmolaridad es la concentración osmolar expresada en osmoles por litro de solución en lugar de osmoles por kilogramo de agua. Aunque en sentido estricto son los osmoles por kilogramo de agua (osmolalidad) los que determinan la presión osmótica, para las soluciones diluidas como las que se encuentran en el cuerpo las diferencias cuantitativas entre la osmolaridad y la osmolalidad son menores del 1%.
Transporte activo: En ocasiones es necesaria una gran concentración de una sustancia en el líquido intracelular aun cuando el líquido extracelular contenga solo una pequeña concentración. Esta situación tiene lugar, por ejemplo, para los iones potasio
Cuando una membrana celular transporta moléculas o iones «contra corriente» contra un gradiente de concentración (o «contra corriente» contra un gradiente eléctrico o de presión), el proceso se denomina transporte activo
En el transporte activo primario la energía procede directamente de la escisión del trifosfato de adenosina (ATP) o de algún otro compuesto de fosfato de alta energía.
El transporte activo secundario la energía procede secundariamente de la energía que se ha almacenado en forma de diferencias de concentración iónica de sustancias moleculares o iónicas secundarias entre los dos lados de una membrana celular, que se generó originalmente mediante transporte activo primario.
En ambos casos el transporte depende de proteínas transportadoras que penetran a través de la membrana celular, al igual que en la difusión facilitada. Sin embargo, en el transporte activo la proteína transportadora funciona de manera diferente al transportador de la difusión facilitada porque es capaz de impartir energía a la sustancia transportada para moverla contra el gradiente electroquímico
los genes, que están situados en el núcleo de todas las células del organismo, controlan la herencia de padres a hijos, pero muchas personas no se dan cuenta de que estos mismos genes también controlan la función cotidiana de todas las células del organismo. Los genes controlan las funciones de la célula
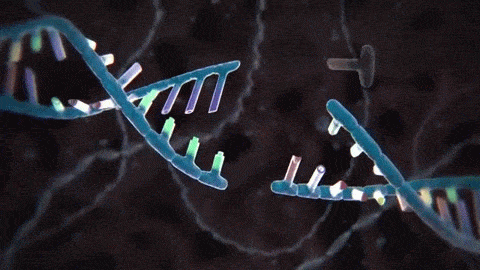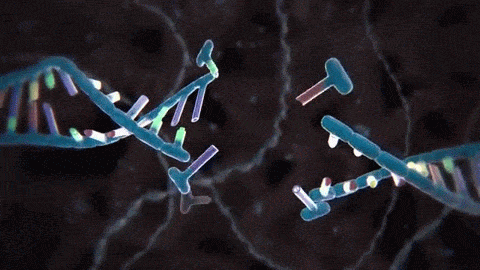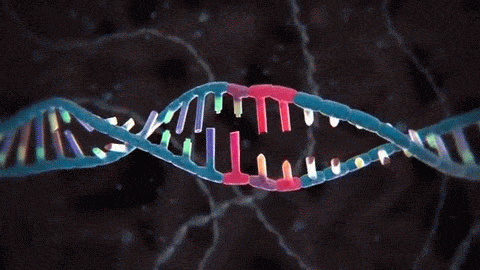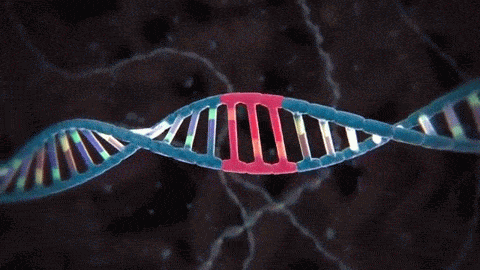
Los genes en el núcleo celular controlan la síntesis de las proteínas: En el núcleo celular hay un gran número de genes unidos por sus extremos, formando las moléculas de doble hélice largas de ADN que tienen un peso molecular que se mide por miles de millones. La molécula está formada por varios compuestos químicos sencillos unidos siguiendo un patrón regular, cuyos detalles pasamos a exponer a continuación.
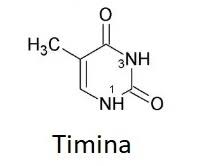
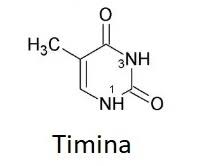
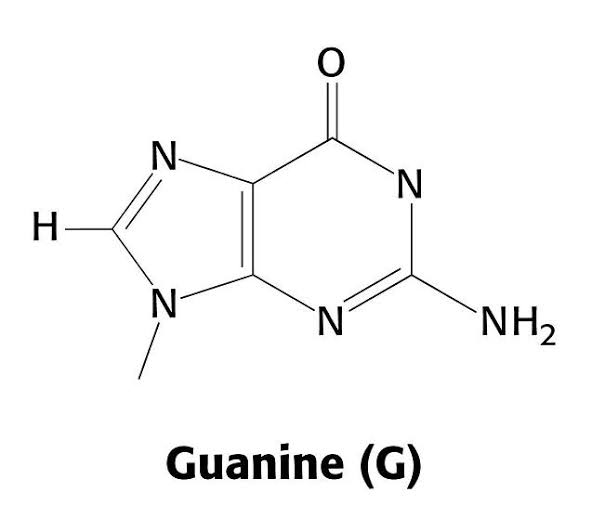
Bloques básicos de ADN: Estos compuestos incluyen: 1) el ácido fosfórico; 2) el azúcar desoxirribosa, y 3) cuatro bases nitrogenadas (dos purínicas, adenina y guanina, y dos pirimidínicas, timina y citosina). El ácido fosfórico y la desoxirribosa forman las dos hebras helicoidales que sirven de soporte para la molécula de ADN
bases nitrogenadas:
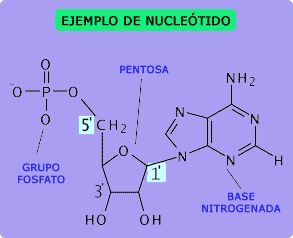
Nucleótidos:
La primera etapa en la formación del ADN consiste en combinar una molécula de ácido fosfórico, una molécula de desoxirribosa y una de las cuatro bases para formar un nucleótido ácido. De esta forma se crean cuatro nucleótidos distintos, uno para cada una de las cuatro bases, los ácidos desoxiadenílico, desoxitimidílico, desoxiguanílico y desoxicitidílico.
El esqueleto de cada hebra de ADN está compuesto por moléculas de ácido fosfórico y desoxirribosa que se van alternando. A su vez, las bases de purina y pirimidina se unen a los lados de las moléculas de desoxirribosa. Después, las dos hebras respectivas de ADN se mantienen unidas mediante enlaces débiles de hidrógeno (líneas discontinuas) entre las bases purínicas y pirimidínicas. La secuencia de los pares de bases complementarios es CG, CG, GC, TA, CG, TA, GC, AT y AT. Debido a la laxitud de los enlaces de hidrógeno, las dos hebras se separan con facilidad y lo hacen muchas veces cuando realizan sus funciones en la célula.
Código genético: La importancia del ADN se debe a su capacidad para controlar la formación de las proteínas en la célula, que se consigue mediante un código genético, Cuando las dos hebras de la molécula de ADN se escinden quedan expuestas las bases purinicas y pirimidinicas a un lado de cada hebra de ADN.
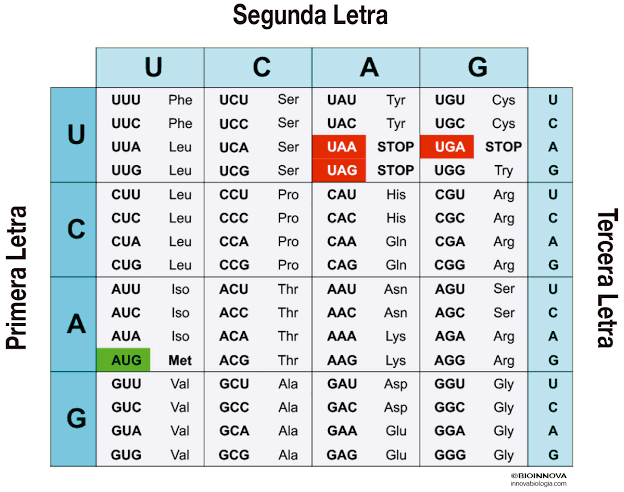
El código genético consta de «tripletes» sucesivos de bases, tres bases sucesivas, componen una palabra del código. Los tripletes sucesivos controlan en último término la secuencia de aminoácidos en una molécula proteica que la célula debe sintetizar.
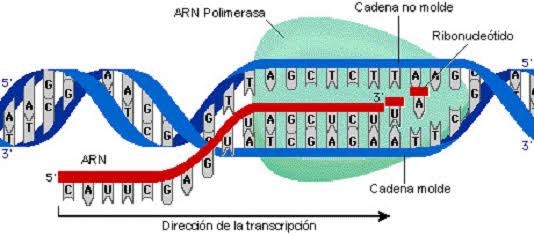
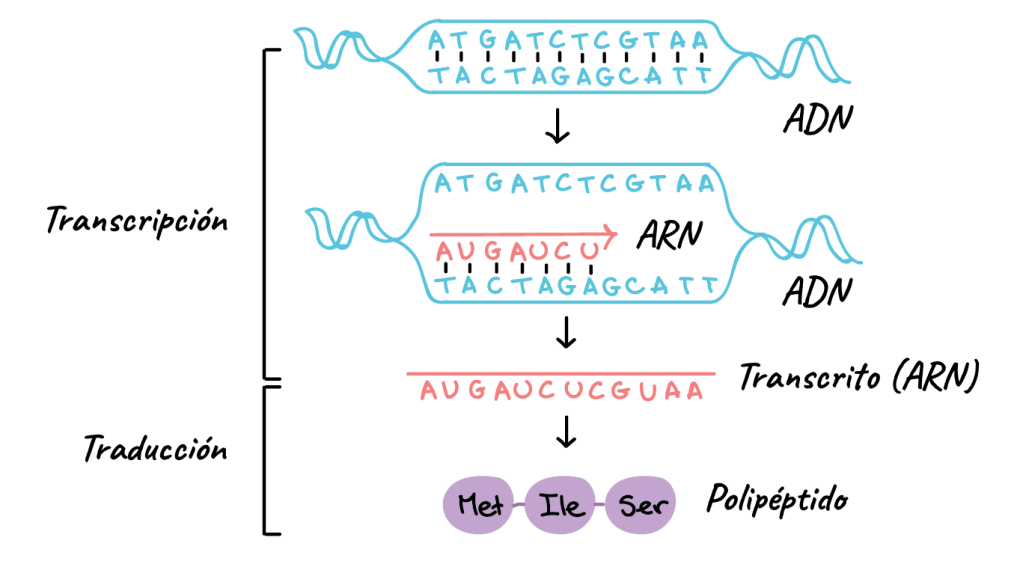
El ARN se sintetiza en el núcleo a partir de una plantilla de ADN: Durante la síntesis de ARN las dos hebras de la molécula de ADN se separan temporalmente y una de ellas se usa como plantilla para la síntesis de una molécula de ARN. Los tripletes del código del ADN provocan la formación de tripletes con un código complementario (o codones) en el ARN. A su vez, estos codones controlarán la secuencia de aminoácidos en una proteína que se va a sintetizar en el citoplasma celular.
ARN mensajero: los codones Las moléculas de ARN mensajero son cadenas largas y sencillas que se encuentran en suspensión en el citoplasma. Estas moléculas están compuestas por varios cientos a miles de nucleótidos de ARN en cadenas no pareadas y contienen codones que son exactamente complementarios a los tripletes del código de los genes de ADN. Sus codones son CCG, UCU y GAA, que son los codones de los aminoácidos prolina, serina y ácido glutámico. «Es el único que codifica para síntesis de proteínas».
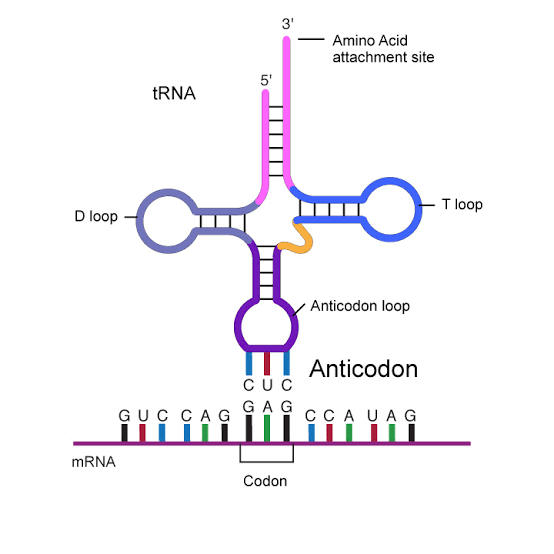
ARN de transferencia: los anticodones Otro tipo de ARN que tiene un papel esencial en la síntesis proteica se conoce como ARN de transferencia (ARNt), porque transfiere las moléculas de aminoácidos a las moléculas proteicas a medida que se va sintetizando la proteína. Cada tipo de ARNt se combina específicamente con 1 de los 20 aminoácidos que se van a incorporar en las proteínas. El ARNt actúa como vehículo para transportar su tipo específico de aminoácido a los ribosomas, donde se van formando las moléculas proteicas., En los ribosomas, cada tipo específico de ARNt reconoce un codón en particular en el ARNm (que se describe más adelante) y libera el aminoácido apropiado en el lugar apropiado de la cadena de la molécula proteica que se está formando.
ARN ribosómico El tercer tipo de ARN de la célula es el ARN ribosómico, que constituye el 60% del ribosoma. El resto está formado por proteínas, unas 75 clases distintas que son de tipo estructural, y también las enzimas necesarias para la fabricación de las moléculas proteicas. El ribosoma es la estructura física del citoplasma en el que se sintetizan realmente las moléculas proteicas. No obstante, siempre funciona asociado a otros tipos de ARN: el ARNt transporta los aminoácidos al ribosoma para su incorporación en la molécula proteica en desarrollo, mientras que el ARNm proporciona la información necesaria para el secuenciado de aminoácidos en el orden
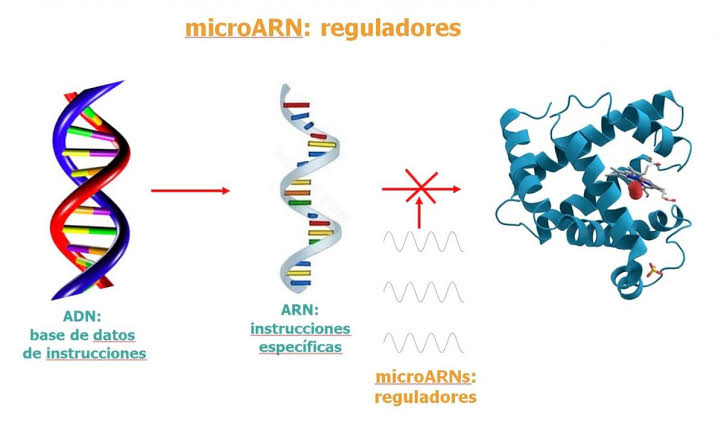
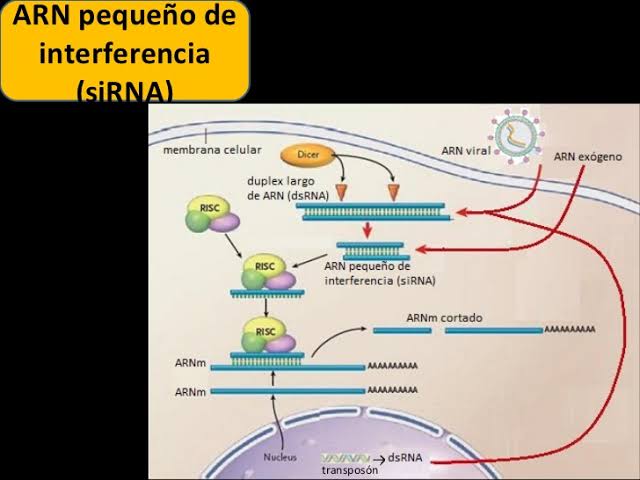
ARNmi y ARN: de interferencia pequeño Un cuarto tipo de ARN en la célula es el microARN (ARNmi), formado por cortos fragmentos de ARN monocatenario (de 21 a 23 nucleótidos) que regulan la expresión génica (fig. 3-10). Los ARNmi se codifican a partir del ADN transcrito de genes, pero no se traducen a proteínas y, por tanto, a menudo reciben el nombre de ARN no codificante. Los ARNmi son procesados por las células en moléculas que son complementarias al ARNm y que actúan para reducir la expresión génica. La generación de ARNmi supone un procesamiento especial de ARN precursores primarios más largos llamados ARNmi-pri, que son los transcriptos primarios del gen.
Formación de proteínas en los ribosomas: el proceso de «traducción» Cuando una molécula de ARNm entra en contacto con un ribosoma, se desplaza por este a partir de un extremo predeterminado de la molécula de ARN que se especifica mediante la secuencia apropiada de las bases de ARN, el codón «iniciador de la cadena». mientras el ARNm se desplaza por el ribosoma, se forma una molécula proteica, en un proceso que se conoce como traducción. Es decir, el ribosoma lee los codones del ARNm igual que una cinta se «lee» cuando va pasando por el cabezal de un magnetófono. Después, se marca el final de una molécula proteica cuando un codón de parada (el codón «terminador de la cadena») atraviesa el ribosoma y la molécula proteica se libera en el citoplasma.
Regulación genética: La regulación genética, o regulación de la expresión génica, cubre todo el proceso, desde la transcripción del código genético en el núcleo hasta la formación de proteínas en el citoplasma. La regulación de la expresión génica dota a todos los organismos vivos de la capacidad para responder a los cambios en su medio
Control de las funciones intracelulares mediante la regulación enzimática Además del control de las funciones de la célula mediante la regulación genética, las actividades celulares están controladas también por inhibidores o activadores intracelulares que actúan directamente sobre las enzimas intracelulares específicas.
Inhibición enzimática Algunas de las sustancias químicas formadas en la célula ejercen una retroalimentación directa inhibiendo los sistemas enzimáticos específicos que los sintetizan. Casi siempre, el producto sintetizado actúa sobre la primera enzima de una secuencia en lugar de hacerlo sobre las enzimas sucesivas, uniéndose directamente a ella y provocando un cambio conformacional alostérico que la inactiva
Activación enzimática Las enzimas que están normalmente inactivas se activan cuando es necesario. Un ejemplo de este fenómeno se produce cuando se ha agotado la mayoría del ATP de la célula. En este caso, comienza a formarse una cantidad considerable de monofosfato de adenosina cíclico (AMPc) como producto de división del ATP.
Ciclo vital de la célula : es el período que transcurre desde el inicio de la reproducción celular hasta el inicio de la siguiente reproducción celular. Cuando las células de los mamíferos no están inhibidas, se reproducen tan rápidamente como pueden y su ciclo vital puede ser tan solo de 10 a 30 h y termina por una serie de sucesos físicos diferenciados que se denominan mitosis y que tienen como finalidad lograr la división de la célula en dos células hijas nuevas, . La mitosis en sí misma dura solo unos 30 min, por lo que más del 95% del ciclo vital de las células está representado por el intervalo entre las mitosis, o interfase, incluso en las células que se reproducen con mayor rapidez.
Control del crecimiento y la reproducción celular: Algunas células crecen y se reproducen continuamente, como las células que dan lugar a los elementos formes sanguíneos en la médula ósea, las capas germinales de la piel y el epitelio intestinal, mientras que muchas otras células, como los miocitos del músculo liso, pueden no reproducirse durante muchos años. Algunas células, como las neuronas y la mayoría de los miocitos del músculo estriado, no se reproducen durante toda la vida de una persona excepto durante el período de vida fetal.
Regulación del tamaño de la célula El tamaño de la célula está determinado casi en su totalidad por la cantidad de ADN funcionante que hay en el núcleo. Si el ADN no se replica, la célula crece hasta un tamaño determinado y después lo mantiene, mientras que también es posible que, usando el quimioterápico colchicina, se impida la formación del huso mitótico y, por tanto, la mitosis aunque continúe la replicación del ADN. En este caso, el núcleo contiene una cantidad de ADN bastante mayor de lo que normalmente contendría y la célula crece hasta un tamaño proporcionalmente mayor. Se supone que este crecimiento celular es consecuencia del aumento de la producción de ARN y de las proteínas celulares, lo que, a su vez, hace que el tamaño de la célula aumente más.
Diferenciación celular Una característica especial del crecimiento y la división celular es la diferenciación celular, que se refiere a los cambios de las propiedades físicas y funcionales de las células a medida que proliferan en el embrión para formar las distintas estructuras y órganos corporales. La siguiente descripción de un experimento especialmente interesante nos permite explicar estos procesos. Cuando el núcleo de una célula de la mucosa intestinal de una rana se implanta quirúrgicamente en el óvulo de una rana en el que se había extraído el núcleo original se consigue crear una rana normal. Este experimento demuestra que incluso una célula de la mucosa intestinal, que es una célula bien diferenciada, contiene toda la información genética necesaria para el desarrollo de todas las estructuras necesarias para el organismo de este animal.
Apoptosis: muerte celular programada Los 100 billones de células del organismo forman parte de una comunidad muy bien organizada en la que el número total de células está regulado no solo por el control de la velocidad de división celular, sino también por el control de la velocidad de la muerte celular. Cuando las células ya no se necesitan, o cuando se convierten en una amenaza para el organismo, sufren una muerte celular programada suicida, o apoptosis. Este proceso implica una cascada proteolítica específica que hace que la célula se encoja y condense para desmontar su citoesqueleto y alterar su superficie de tal forma que una célula fagocítica cercana, como un macrófago, se pueda unir a la membrana celular y digerir la célula.
Cáncer: El cáncer se debe en la mayoría de los casos a la mutación o a alguna otra activación anormal de los genes celulares que controlan el crecimiento y la mitosis celular. Los protooncogenes son genes normales que codifican diversas proteínas responsables del control de la adhesión celular, el crecimiento y la visión. Si mutan o se activan de forma excesiva, los protooncogenes pueden convertirse en oncogenes con funcionamiento anómalo capaces de provocar cáncer. En los cánceres humanos se han descubierto hasta 100 tipos distintos de oncogenes. En todas las células también hay antioncogenes, denominados también genes supresores de tumores, que suprimen la activación de los oncogenes específicos, es decir, la pérdida o inactivación de los antioncogenes permite la activación de los oncogenes que conduce al cáncer
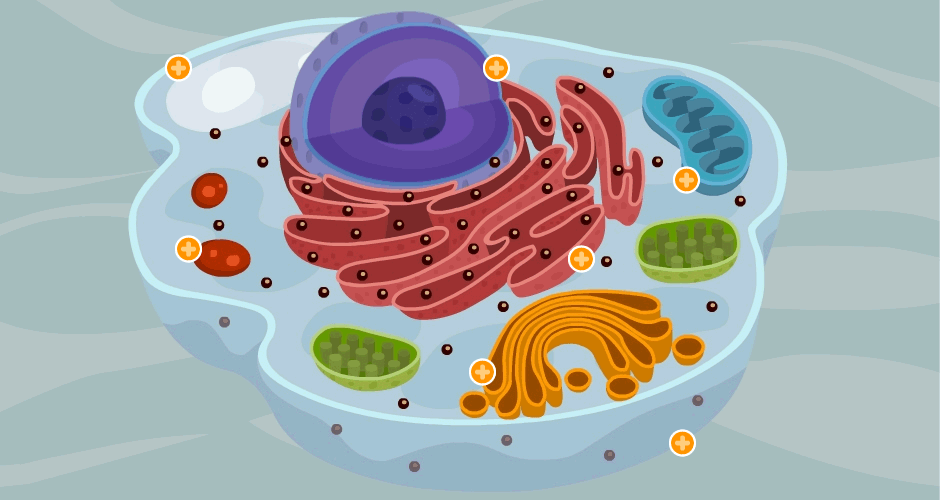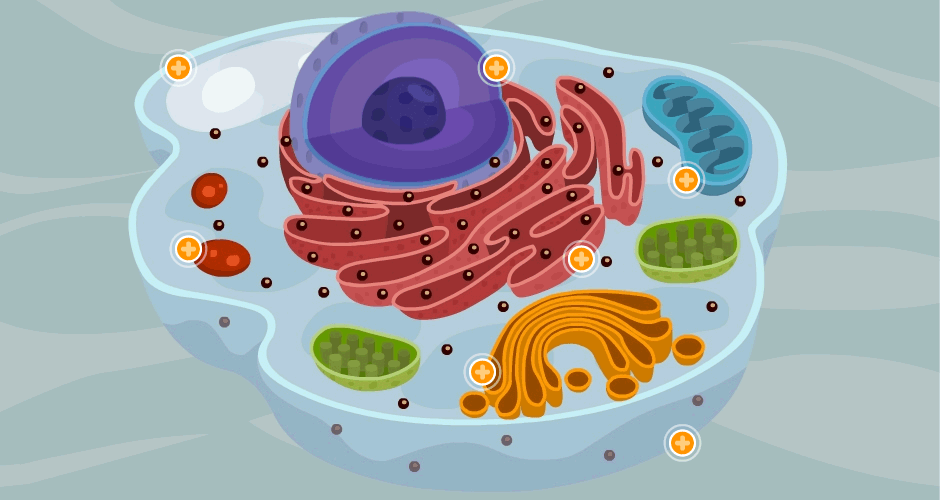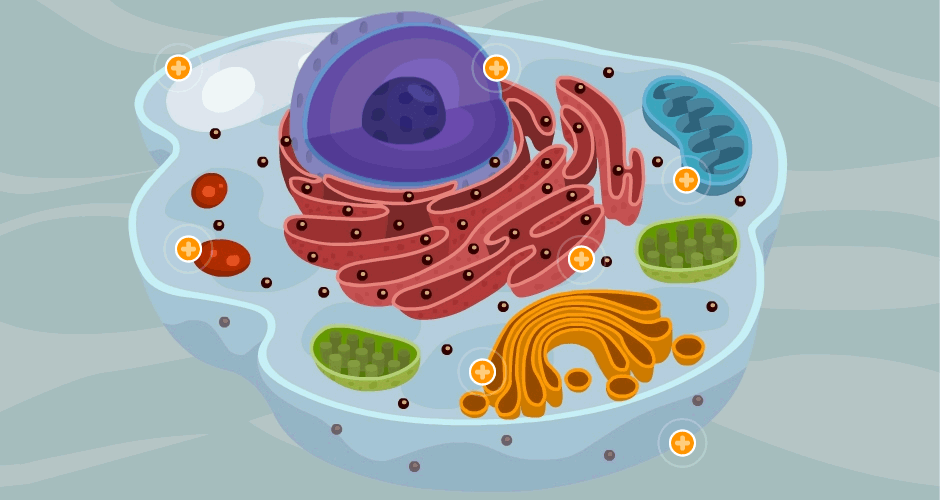
Las células son los elementos básicos que conforman el organismo, ingieren los nutrientes y los convierten en energía, y realizan funciones especializadas. Contienen el código hereditario del organismo que controla las sustancias sintetizadas por las células y les permite realizar copias de sí mismas.
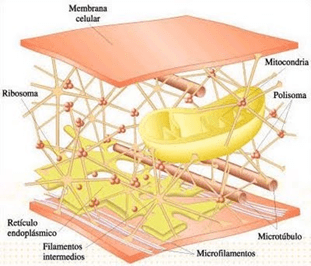
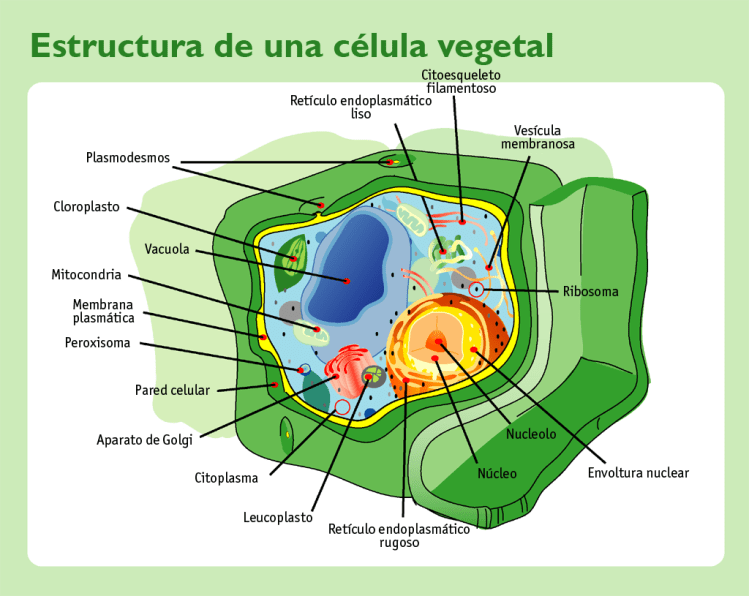
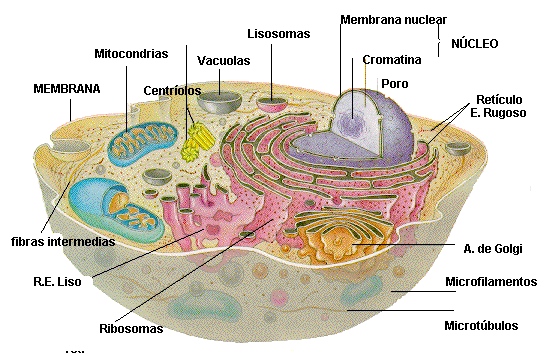
Sus dos partes más importantes son el núcleo y el citoplasma, que están separados entre sí por una membrana nuclear.
Sustancias que componen la célula se conocen colectivamente como protoplasma.
La célula contiene estructuras físicas muy organizadas que se denominan orgánulos intracelulares.
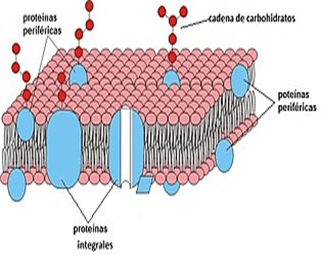
Membrana celular: Es una estructura elástica, fina y flexible que tiene un grosor de tan solo 7,5 a 10 nm. una composición aproximada de un 55% de proteínas, un 25% de fosfolípidos, un 13% de colesterol, un 4% de otros lípidos y un 3% de hidratos de carbono, La barrera lipídica de la membrana celular impide la penetración de sustancias hidrosolubles.
Proteínas integrales y periféricas: Estas proteínas de membrana son principalmente glucoproteínas, proteínas integrales que protruyen por toda la membrana y proteínas periféricas que se unen solo a una superficie de la membrana y que no penetran en todo su espesor, Muchas de las proteínas integrales componen canales estructurales (o poros) a través de los cuales las moléculas de agua y las sustancias hidrosolubles, especialmente los iones, pueden difundir entre los líquidos extracelular e intracelular.
Hidratos de carbono de la membrana: «glucocáliz» celular
Los hidratos de carbono de la membrana se presentan casi invariablemente combinados con proteínas o lípidos enforma de glucoproteínas o glucolípidos. Toda la superficie externa de la célula a menudo contiene un recubrimiento débil de hidratos de carbono que se conoce como glucocáliz.
Citoplasma y sus orgánulos: El citoplasma está lleno de partículas diminutas y grandes y orgánulos dispersos……
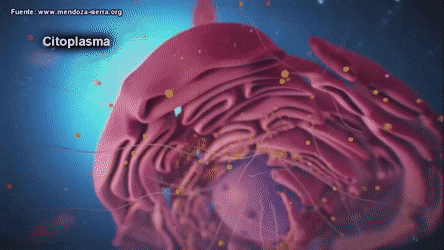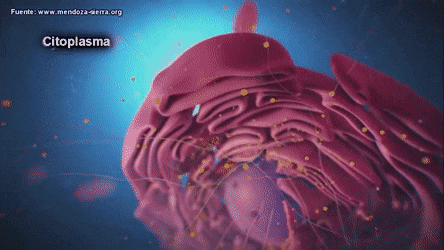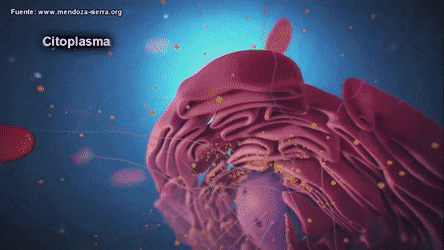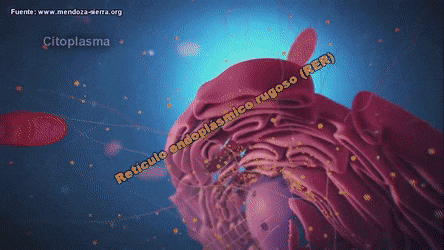
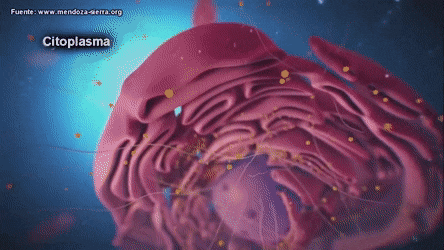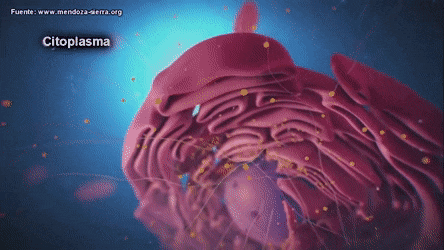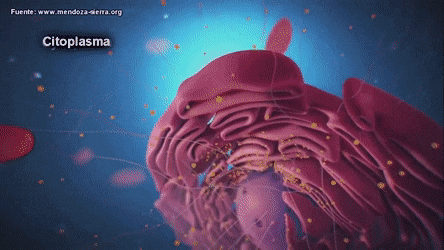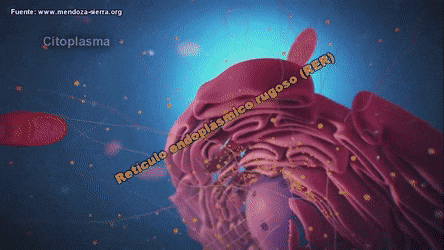
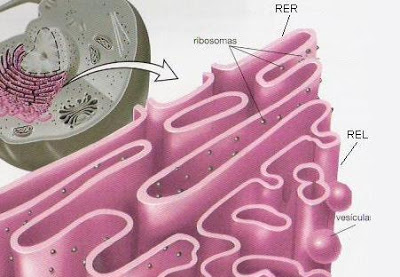
ribosomas y retículo endoplásmico rugoso
retículo endoplásmico liso o agranular
Unidas a la superficie exterior de muchas partes del retículo endoplásmico encontramos una gran cantidad de partículas granulares diminutas que se conocen como ribosomas. Los ribosomas están formados por una mezcla de ARN y proteínas y su función consiste en sintetizar nuevas moléculas proteicas en la célula.
Este retículo agranular actúa en la síntesis de sustancias lipídicas y en otros procesos de las células que son promovidos por las enzimas intrarreticulares
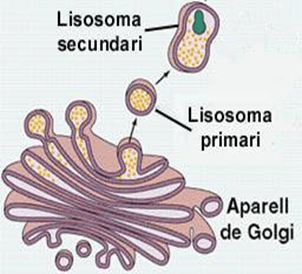
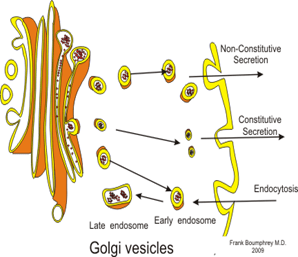
Aparato de Golgi:
Está íntimamente relacionado con el retículo endoplásmico. Tiene unas membranas similares a las del retículo endoplásmico a granular y está formado habitualmente por cuatro o más capas apiladas de vesículas cerradas, finas y planas, que se alinean cerca de uno de los lados del núcleo.
Lisosomas:
Son orgánulos vesiculares que se forman por la rotura del aparato de Golgi y después se dispersan por todo el citoplasma. Los lisosomas constituyen el aparato digestivo intracelular que permite que la célula digiera:
Estructuras dañadas
Partículas del alimento
Sustancias no deseadas como bacterias.
El lisosoma es muy distinto en los diferentes tipos celulares, pero habitualmente tiene un diámetro de 250 a 750 nm.
Peroxisomas:
Se cree que están formados por autorreplicación (o, quizás, protruyendo desde el retículo endoplásmico liso) en lugar de proceder del aparato de Golgi. Contienen oxidasas en lugar de hidrolasas, El peróxido de hidrógeno es una sustancia muy oxidante que actúa junto con una catalasa, otra enzima oxidasa que se encuentra en grandes cantidades en los peroxisomas para oxidar muchas sustancias que, de lo contrario, serían venenosas para la célula.
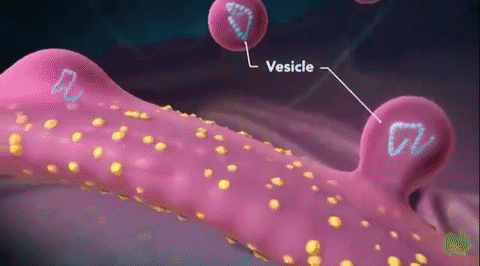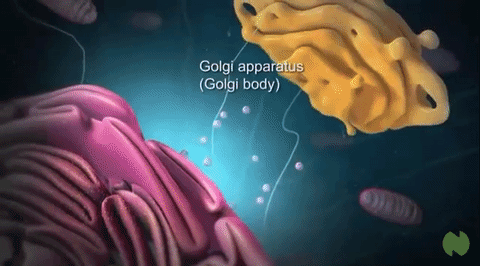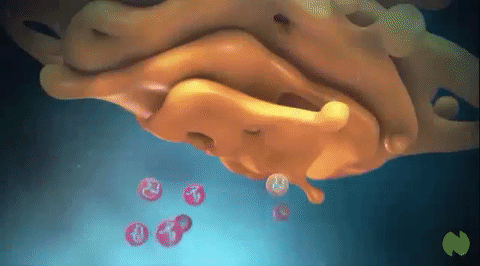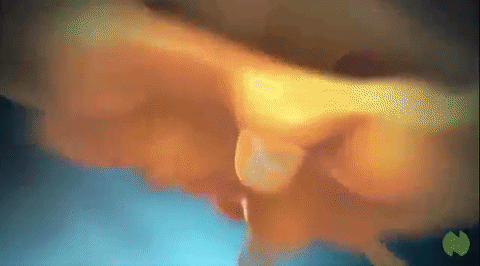
Vesículas secretoras: Casi todas las sustancias secretoras se forman en el sistema retículo endoplásmico-aparato de Golgi y después se liberan desde el aparato de Golgi hacia el citoplasma en forma de vesículas de almacenamiento que se conocen como vesículas secretoras o gránulos secretores.
Estas vesículas almacenan proenzimas proteicas (enzimas que aún no están activadas) que se segregan más tarde a través de la membrana celular hacia el conducto pancreático, es decir, hacia el duodeno, donde se activan y realizan sus funciones digestivas sobre el alimento en el aparato digestivo.
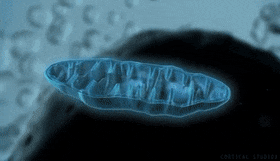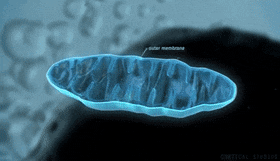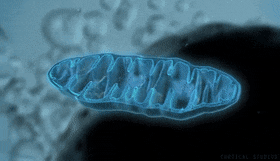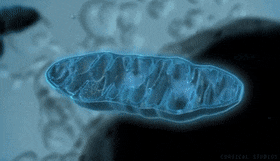
Mitocondrias: Se conocen como los «centros neurálgicos» de la célula. Sin ellas, las células no serían capaces de extraer energía suficiente de los nutrientes y, en esencia, cesarían todas las funciones celulares. Las mitocondrias se encuentran en todas las zonas del citoplasma de la célula, pero su número total en cada célula varía de menos de cien hasta varios miles, dependiendo de la cantidad de energía que requiere la célula. Algunas mitocondrias miden solo algunos cientos de nanómetros de diámetro y adoptan forma globular, mientras que otras son alargadas, miden hasta 1 μm de diámetro y 7 μm de longitud.
Citoesqueleto celular:
El citoesqueleto celular es una red de proteínas fibrilares organizadas habitualmente en filamentos o túbulos que se originan como moléculas proteicas precursoras sintetizadas por los ribosomas en el citoplasma, para formar un soporte elástico para la membrana celular. Todas las células usan un tipo especial de filamento rígido formado por polímeros de tubulina para construir estructuras tubulares fuertes, los microtúbulos.
Núcleo:
El núcleo, que es el centro de control de la célula, envía mensajes a esta para que crezca y madure, se replique o muera.Contiene grandes cantidades de ADN, que comprende los genes, que son los que determinan las características de las proteínas celulares, como las proteínas estructurales, y también las enzimas intracelulares que controlan las actividades citoplásmicas y nucleares. Los genes también controlan y promueven la reproducción de la célula. Los genes se reproducen primero para crear dos juegos idénticos de genes y después se divide la célula utilizando un proceso especial, que se conoce como mitosis, para formar dos células hijas, cada una de las cuales recibe uno de los dos juegos de genes de ADN.
Comparación entre la célula animal y las formas de vida precelulares:
La célula es un organismo complicado que ha necesitado muchos cientos de millones de años para desarrollarse después de que apareciera la primera forma de vida, un organismo similar a los virus de nuestros días, sobre la tierra.
Tamaños relativos de:
1) el virus más pequeño conocido;
2) un virus grande;
3) una rickettsia;
4) una bacteria,
5) una célula nucleada, donde se ve que la célula tiene un diámetro en torno a 1.000 veces mayor que el del virus más pequeño y, por tanto, un volumen en torno a 1.000 millones de veces mayor que el del virus más pequeño.
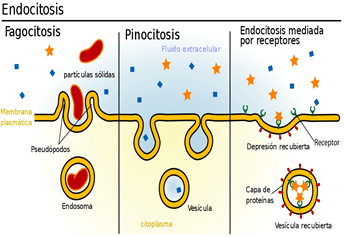
pinocitosis se produce continuamente en las membranas celulares de la mayoría de las células, pero es especialmente rápida en algunas de ellas. Por ejemplo, es muy rápida en los macrófagos, donde aproximadamente el 3% del total de su membrana es engullido en forma de vesículas cada minuto. las vesículas de pinocitosis son tan pequeñas, habitualmente de solo 100 a 200 nm de diámetro, que la mayoría de ellas solo se pueden ver con un microscopio electrónico.
Fagocitosis:
Solo algunas células tienen la capacidad de realizar la fagocitosis, principalmente los macrófagos tisulares y algunos leucocitos sanguíneos. La fagocitosis se inicia cuando una partícula, como una bacteria, una célula muerta o un resto de tejido, se une a los receptores de la superficie de los fagocitos.
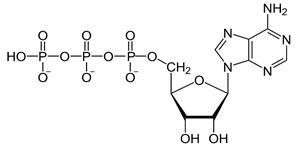
Características funcionales del ATP
El ATP es un nucleótido compuesto por:
1) la base nitrogenada adenina
2) el azúcar pentosa ribosa
3) tres radicales fosfato
La energía liberada se usa para dar energía a muchas de las demás funciones celulares, como la síntesis de sustancias y la contracción muscular. El ATP se conoce como la moneda energética de la célula porque se puede gastar y recomponer continuamente, con un ciclo metabólico de solo unos minutos. La energía del ATP se usa para promover tres categorías principales de funciones celulares.
Transporte de sustancias a través de múltiples membranas en la célula;
Síntesis de compuestos químicos a través de la célula
Trabajo mecánico.
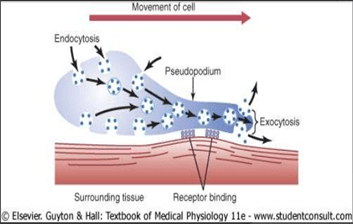
Movimiento amebiano:
El movimiento amebiano es el movimiento de toda la célula en relación con su entorno, como el movimiento de los leucocitos a través de los tejidos. El movimiento amebiano comienza con la protrusión de un seudópodo desde un extremo de la célula. Este seudópodo se proyecta lejos de la célula y se asegura parcialmente en una zona nueva.
Cilios:
El movimiento ciliar, es un movimiento a modo de látigo de los cilios que se encuentran en la superficie de las células. Este movimiento existe principalmente solo en dos lugares del cuerpo humano: en la superficie de las vías aéreas y en la superficie interna de las trompas uterinas (trompas de Falopio) del aparato reproductor.
El movimiento de látigo de los cilios de la cavidad nasal y las vías aéreas bajas hace que una capa de moco se desplace a una velocidad aproximada de 1 cm/min hacia la faringe, con lo que el moco y las partículas que han quedado atrapadas en el moco de estos conductos se están limpiando continuamente. El movimiento del cilio, que se desplaza hacia delante con un movimiento rápido, como un golpe de látigo, con una frecuencia de 10 a 20 veces por segundo, doblándose bruscamente en el punto en el que se proyecta desde la superficie de la célula.
El flagelo de un espermatozoide es similar a un cilio; tiene el mismo tipo de estructura y el mismo tipo de mecanismo contráctil. Este flagelo es mucho más largo y se desplaza con ondas de tipo cuasi-sinusoidal en lugar de movimientos de tipo látigo.
La fisiología es la ciencia que pretende explicar los mecanismos físicos y químicos responsables del origen, desarrollo y progresión de la vida. Desde el virus más simple hasta el árbol más grande o el complicado ser humano.
Fisiología vírica,
Fisiología bacteriana,
fisiología celular,
fisiología vegetal
fisiología de los invertebrados
fisiología de los vertebrados
fisiología de los mamíferos
Fisiología humana.
La unidad viva básica del cuerpo es la célula. Aunque los eritrocitos son las más abundantes entre todas las células corporales, hay 75 billones de células más de otros tipos que realizan funciones diferentes. El cuerpo en su conjunto contiene en torno a 100 billones de células.
Liquido extracelular e intracelular:
El 60% del cuerpo humano del adulto es líquido, principalmente una solución acuosa de iones y otras sustancias. Este líquido queda dentro de las células y se conoce como líquido intracelular, El líquido extracelular está en movimiento constante por todo el cuerpo y se transporta rápidamente en la sangre circulante
Hemostasis: mantenimiento de un medio interno casi constante.
En 1929, el fisiólogo estadounidense Walter Cannon (1871-1945) acuñó el término homeostasis para referirse al mantenimiento de unas condiciones casi constantes del medio interno.
La enfermedad se considera un estado de ruptura de la homeostasis. Sin embargo, incluso en presencia de enfermedades, los mecanismos homeostáticos siguen activos y mantienen las funciones vitales a través de múltiples compensaciones.
El aparato circulatorio:
El líquido extracelular circula por el organismo en dos etapas. La primera de ellas consiste en el movimiento de la sangre por el cuerpo dentro de los vasos sanguíneos, y la segunda es el movimiento del líquido entre los capilares sanguíneos y los espacios intercelulares entre las células tisulares.
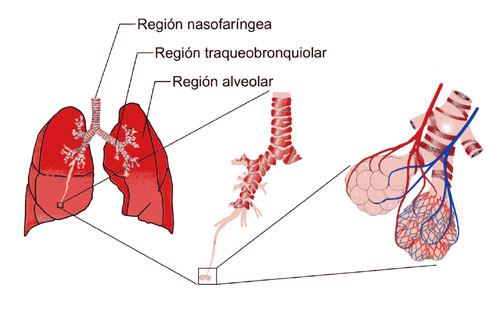
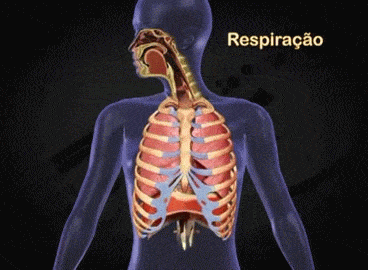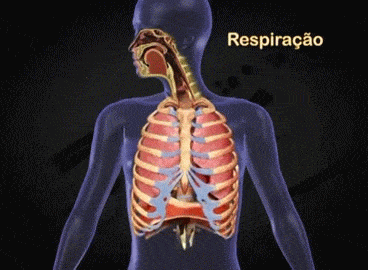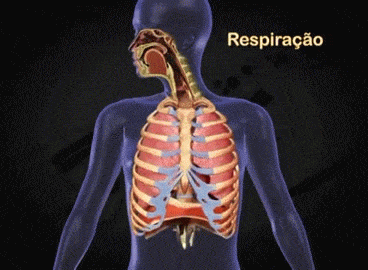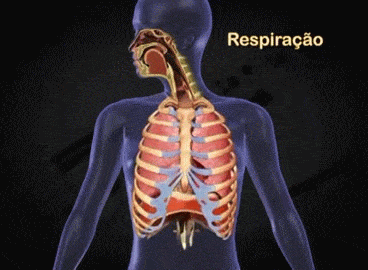
Aparato respiratorio: Cada vez que la sangre atraviesa el organismo también fluye por los pulmones y capta el oxígeno a través de los alvéolos, la membrana alveolar, tiene un grosor de tan solo 0,4 a 2 μm y el oxígeno difunde rápidamente por el movimiento molecular.
Aparato digestivo:
Una gran porción de la sangre que bombea el corazón también atraviesa las paredes del aparato digestivo, donde se absorben los distintos nutrientes, incluidos los hidratos de carbono, los ácidos grasos y los aminoácidos, desde el alimento ingerido hacia el líquido extracelular de la sangre. El hígado es el encargado de cambiar la composición química de muchas de las sustancias absorbidas. Entre las funciones del hígado se encuentra la detoxificación o eliminación de numerosos fármacos y productos químicos que se ingieren.
Regulación de las funciones corporales:
El sistema nervioso está compuesto por tres partes principales: la porción de aferencia sensitiva, el sistema nervioso central (o la porción integradora) y la porción eferente motora. Los receptores sensitivos detectan el estado del cuerpo o de su entorno, Un segmento importante del sistema nervioso es el sistema nervioso autónomo o neurovegetativo, que funciona a escala subconsciente y controla muchas de las funciones de los órganos internos, como la función de bomba del corazón, los movimientos del aparato digestivo y la secreción en muchas de las glándulas corporales.
Sistemas hormonales:
La hormona tiroidea aumenta la velocidad de la mayoría de las reacciones químicas de todas las células.
La insulina controla el metabolismo de la glucosa.
Las hormonas corticosuprarrenales controlan los iones sodio y potasio y el metabolismo proteico.
La hormona paratiroidea controla el calcio y el fosfato en el hueso.
Las hormonas se transportan en el líquido extracelular a otras partes del cuerpo para regular las funciones celulares.
El sistema inmunitario:
está formado por los glóbulos blancos, células tisulares derivadas de los glóbulos blancos, el timo, los nódulos linfáticos y los vasos linfáticos que protegen el cuerpo de patógenos como bacterias, virus, parásitos y hongos. diferencia sus propias células de las células y sustancias extrañas.
Sistema tegumentario:
La piel y sus diversos anejos, como el pelo, las uñas, las glándulas etc. cubren, amortiguan y protegen los tejidos profundos y los órganos del cuerpo.
El sistema tegumentario es importante también para la regulación de la temperatura y la excreción de los residuos.
La piel suele comprender entre aproximadamente el 12 y el 15% del peso corporal, es importante también para la regulación de la temperatura y la excreción de los residuos.
Reproducción:
No se considera que la reproducción sea una función homeostática, aunque ayuda a mantener la homeostasis generando nuevos seres que ocuparán el lugar de aquellos que mueren.
Sistemas de control del organismo:
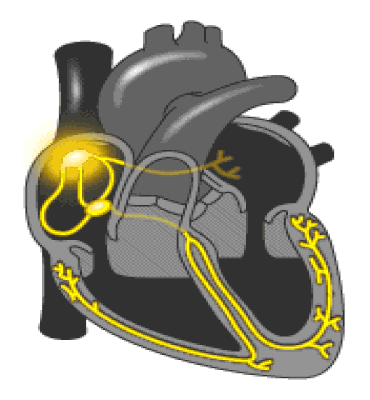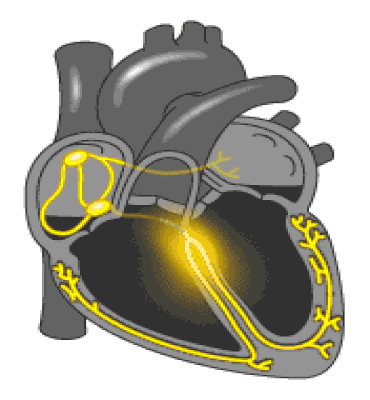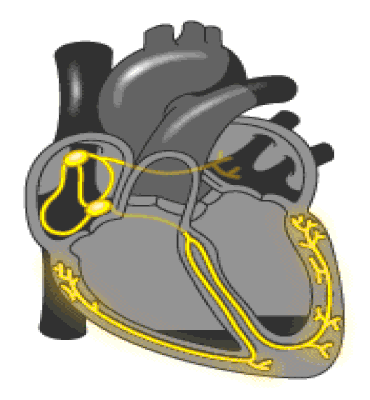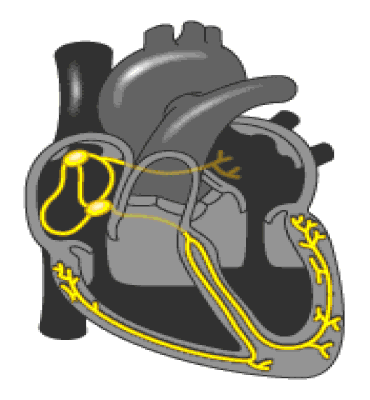
Regulación de la presión arterial: El sistema de barorreceptores; En las paredes de la zona en que se bifurcan las arterias carótidas en el cuello, y también en el cayado aórtico en el tórax, se encuentran muchos receptores nerviosos denominados barorreceptores, envían descargas de impulsos nerviosos al bulbo raquídeo cerebral, que es donde estos impulsos inhiben el centro vasomotor y disminuyen el número de impulsos transmitidos desde el centro vasomotor.
Retroalimentación negativa;
La concentración elevada de dióxido de carbono inicia una serie de sucesos que disminuyen la concentración hacia la normalidad, lo que es una señal negativa para iniciar el estímulo.
Esta es la primera entrada de mi nuevo blog. Acabo de empezar a escribir este nuevo blog. ¡Mantente al día para leer más entradas! Suscríbete a continuación para recibir notificaciones cuando publique nuevo contenido.
Esto es una entrada de muestra, originalmente publicada como parte de Blogging University. Regístrate en uno de nuestros diez programas y empieza tu blog con buen pie.
Hoy vas a publicar una entrada. No te preocupes por el aspecto de tu blog. Tampoco te preocupes si todavía no le has puesto un nombre o si todo esto te agobia un poco. Tan solo haz clic en el botón «Nueva entrada» y explícanos por qué estás aquí.
¿Por qué es necesario?
Porque proporciona contexto a los nuevos lectores. ¿A qué te dedicas? ¿Por qué deberían leer tu blog?
Porque te ayudará a concentrarte en tus propias ideas sobre tu blog y en lo que quieres hacer con él.
La entrada puede ser corta o larga, una introducción de tu vida o una declaración de los objetivos del blog, un manifiesto de cara al futuro o una breve descripción del tipo de cosas que pretendes publicar.
Te ofrecemos algunos consejos útiles para ayudarte a empezar:
¿Por qué publicas tus entradas en lugar de escribir en un diario personal?
¿Sobre qué temas crees que escribirás?
¿Con quién te gustaría conectar a través de tu blog?
Si tu blog resulta ser un éxito a lo largo del próximo año, ¿qué objetivo te gustaría lograr?
No tienes por qué atarte a las decisiones que tomes ahora. Lo bueno de los blogs es que evolucionan constantemente a medida que aprendemos nuevas cosas, crecemos e interactuamos los unos con los otros. Pero está bien saber dónde y por qué empezaste, y plasmar tus objetivos puede darte más ideas para las entradas que quieres publicar.
¿No sabes por dónde empezar? Tan solo escribe lo primero que se te ocurra. Anne Lamott, autora de un libro sobre cómo escribir que nos encanta, afirma que debemos permitirnos escribir un «primer borrador de mierda». Anne está en lo cierto: tan solo tienes que empezar a escribir, y ya te encargarás de editarlo más tarde.
Cuando todo esté listo para publicarse, asigna entre tres y cinco etiquetas a la entrada que describan el centro de atención de tu blog: escritura, fotografía, ficción, educación, comida, coches, películas, deportes… ¡Lo que sea! Estas etiquetas ayudarán a los usuarios interesados en tus temas a encontrarte en el Lector. Una de las etiquetas debe ser «zerotohero», para que los nuevos blogueros también puedan encontrarte.